Selenium — Benefits Deep Dive
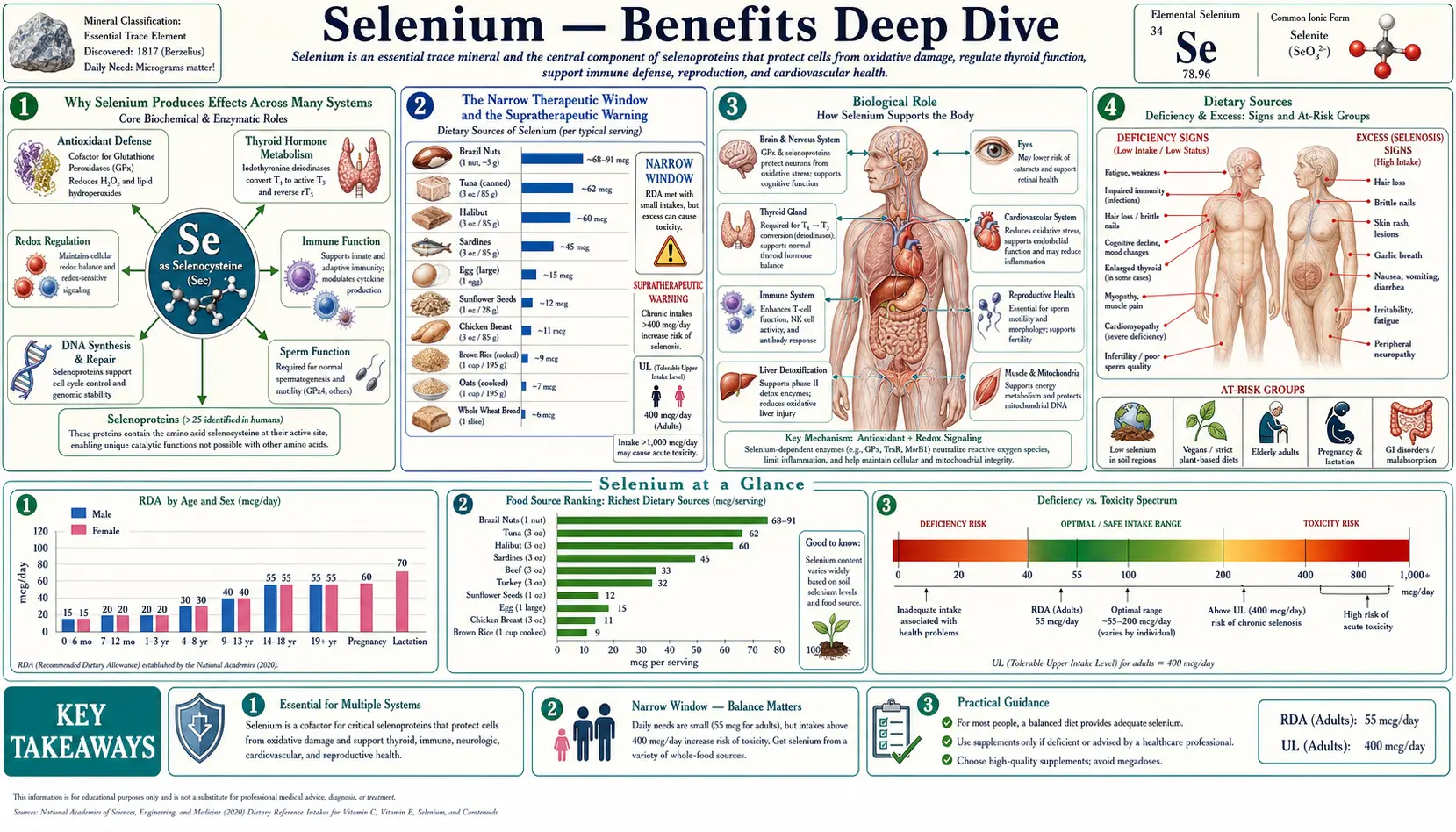
Selenium is a trace element with the narrowest therapeutic window of any essential nutrient. The adult RDA is just 55 micrograms per day; the tolerable upper intake level is 400 micrograms per day. That seven-fold gap between requirement and toxicity is far smaller than for vitamin C (~250-fold), zinc (~20-fold), or magnesium (~10-fold). A single Brazil nut harvested from selenium-rich Brazilian soil can contain anywhere from 5 to 100+ micrograms — eating 6 to 10 per day for months has produced documented cases of selenosis. Within that narrow window, however, selenium is indispensable. It is the only trace element with its own dedicated amino acid (selenocysteine, the 21st amino acid) and its own dedicated co-translational insertion machinery. The four deep-dive pages below explore the four clinical territories where selenium produces the largest, best-characterized effects — cellular antioxidant defense via the selenoprotein family, autoimmune and synthetic regulation of thyroid hormone, the unusual immune effects that include preventing viral mutation and the SELECT prostate-cancer signal, and the prostate-cancer-prevention story whose NPC enthusiasm was substantially tempered by SELECT.
Deep-Dive Articles
Antioxidant Defense
The eight glutathione peroxidase isoforms (GPx1–8) each guarding a different cellular compartment, the three thioredoxin reductases (TrxR1–3), methionine sulfoxide reductase B1, selenoprotein P as the brain-and-testis selenium transport vehicle, the strange UGA-codon read-through translational machinery that makes selenocysteine the 21st amino acid, the dietary selenoamino acids (selenomethionine, selenocysteine, methylselenocysteine), Brazil-nut soil variability that drives selenosis, and the marine selenoneine that concentrates in tuna mitochondria.
Immune Function
The Keshan disease / Coxsackievirus B3 story — how selenium deficiency in the host selects for more virulent viral variants, the six fixed point mutations that converted a benign CVB3 strain into the Keshan cardiotoxin, T-cell proliferation and Th1/Th2 balance, macrophage M1/M2 polarization, NK cell cytotoxicity, the HIV CD4 / selenium relationship, influenza severity, the controversial SISPCT high-dose IV selenium sepsis trial, and the SELECT high-grade prostate cancer signal that has reframed adult selenium supplementation.
Thyroid Function
The thyroid contains more selenium per gram of tissue than any other organ. Iodothyronine deiodinases (DIO1, DIO2, DIO3) activate and inactivate thyroid hormone; glutathione peroxidases (especially GPx3 in the follicular lumen) quench the H2O2 generated during thyroid hormone synthesis; selenium repletion lowers anti-TPO antibodies by 20–40% in Hashimoto's, is first-line therapy for mild Graves' orbitopathy per EUGOGO, and reduces postpartum thyroiditis in anti-TPO-positive women.
Cancer Prevention
Antioxidant protection of DNA from oxidative mutagenesis, p53 / BRCA1 / Gadd45 modulation, selective apoptosis induction in transformed cells by methylselenol and selenodiglutathione, NK and CTL surveillance enhancement, the seminal 1996 NPC trial reporting 52% prostate cancer reduction in low-baseline men, and the SELECT trial null result with high-grade prostate cancer signal that reframed the field around a U-shaped dose-response curve.
Table of Contents
- Deep-Dive Articles
- Why Selenium Produces Effects Across Many Systems
- The Narrow Therapeutic Window and the Supratherapeutic Warning
- Research Papers: Antioxidant Defense and Selenoproteins
- Research Papers: Thyroid Function
- Research Papers: Immune Function and Viral Mutation
- Research Papers: Cancer Prevention (NPC, SELECT)
- Research Papers: Cross-Cutting (Status, Toxicity, Form)
- External Authoritative Resources
- Connections
- Featured Videos
Why Selenium Produces Effects Across Many Systems
Most trace elements act through one principal class of enzymes (zinc through its hundreds of metalloenzymes; iron through hemoproteins and iron-sulfur clusters; copper through its handful of cuproenzymes). Selenium is different because it acts almost exclusively through one structural strategy — the incorporation of selenocysteine at the active site of 25 human selenoproteins — but the resulting enzymes serve three fundamentally different cellular jobs, each of which produces a distinct category of clinical effect.
- Redox defense (the GPx and TrxR families) — The selenocysteine selenol group is a far more efficient peroxide-reducing catalyst than the cysteine thiol because of its lower pKa, larger atomic radius, and more reversible oxidation states. The glutathione peroxidases (GPx1-8) and thioredoxin reductases (TrxR1-3) are the cell's most efficient redox-buffering enzymes, and they are the cellular antioxidant defenses that operate at the catalytic-enzyme level rather than the sacrificial-small-molecule level (vitamins C, E, glutathione). When the GPx/TrxR network is compromised, downstream consequences include lipid peroxidation in membranes (ferroptosis, the master regulator of which is GPx4), accumulation of mutagenic 8-oxoguanine in DNA, oxidative inactivation of redox-sensitive transcription factors, and elevated viral mutation rates in infected cells — the foundation of the Antioxidant Defense page.
- Thyroid hormone metabolism (the deiodinase family DIO1, DIO2, DIO3) — The iodothyronine deiodinases use selenocysteine at the active site to catalyze the regiospecific removal of iodine atoms from thyroxine, either activating it to triiodothyronine (T3) or inactivating it to reverse T3. Selenium is therefore the rate-limiting input for thyroid hormone activation in peripheral tissues. The thyroid gland itself contains more selenium per gram than any other organ, both because of the deiodinases and because GPx3 in the follicular lumen quenches the hydrogen peroxide generated during thyroid hormone synthesis. The full clinical picture is on the Thyroid Function page.
- Selective metabolite-mediated effects (methylselenol, selenodiglutathione) — A subset of dietary selenium compounds (specifically methylselenocysteine from selenium-enriched garlic and broccoli) is metabolized to methylselenol, a small lipophilic selenium species that selectively induces apoptosis in transformed cells, inhibits angiogenesis, and modulates the tumor microenvironment. This is a fundamentally different mechanism from the selenoenzyme-mediated effects above, and it is the mechanistic basis for the chemopreventive properties explored on the Cancer Prevention page.
The three mechanisms together explain why selenium deficiency produces such heterogeneous syndromes — Keshan cardiomyopathy from compromised cardiac GPx and TrxR activity, Hashimoto's and Graves' orbitopathy from compromised thyroidal GPx3 and immune dysregulation, increased high-grade prostate cancer from the loss of selective methylselenol-driven apoptosis of transformed cells in the prostate. The same heterogeneity explains why supratherapeutic selenium is also problematic: pushing already-replete cells beyond saturation does not improve selenoenzyme activity (which is already at maximum) and may produce off-target effects through non-specific incorporation of selenomethionine into the methionine pool.
The Narrow Therapeutic Window and the Supratherapeutic Warning
This hub page deviates from the standard structure to spend a paragraph on a warning that applies across all four deep-dive topics: selenium has the narrowest therapeutic window of any essential nutrient, and the evidence that more is better evaporates rapidly once baseline selenium status is adequate.
- The 1996 Nutritional Prevention of Cancer (NPC) trial reported a 52% reduction in prostate cancer incidence at 200 mcg/day selenized yeast in men with low baseline selenium. That single secondary-endpoint finding launched a decade of enthusiastic selenium supplementation.
- The 2008 Selenium and Vitamin E Cancer Prevention Trial (SELECT), 35,533 men, found no overall prostate cancer benefit from 200 mcg/day selenomethionine and was stopped early. Subsequent analysis found that men in the top baseline-selenium quartile (plasma selenium >137 ng/mL) experienced a 91% increase in high-grade prostate cancer with supplementation.
- The long-term follow-up of NPC participants showed a small but significant increase in incident type 2 diabetes in the 200 mcg/day arm among already-replete participants.
- Multiple meta-analyses now find U-shaped relationships between plasma selenium and all-cause mortality, cardiovascular mortality, and several cancer endpoints, with the "sweet spot" roughly 90 to 140 ng/mL plasma selenium. Below ~70 ng/mL is deficient; above ~140 to 150 ng/mL is supratherapeutic and likely net-harmful.
- A single Brazil nut from selenium-rich Brazilian soil can contain anywhere from 5 to 100+ mcg of selenium, with content driven almost entirely by the soil where the tree grew. Eating 6 to 10 Brazil nuts per day is the most reasonably common way for an American adult to accidentally cross into selenosis territory.
The practical translation: assess selenium status before supplementing. The simplest screening tool is plasma selenium (target 90 to 140 ng/mL). A more sensitive marker is plasma selenoprotein P (SELENOP, target 70 to 90 mcg/g). Supplement to a target, not beyond it. The 200 mcg/day doses used in the Hashimoto's, Graves' orbitopathy, and (in deficient populations) cancer-prevention trials are appropriate for low-baseline adults and time-limited use; they are not appropriate as indefinite "wellness" doses in replete adults. The same supplement that lowered anti-TPO in deficient Hashimoto's patients increased high-grade prostate cancer in replete SELECT participants. There is no "selenium for general antioxidant support" recommendation; there are specific indications backed by specific evidence in specific populations.
Research Papers: Antioxidant Defense and Selenoproteins
- Labunskyy VM, Hatfield DL, Gladyshev VN (2014). Selenoproteins: molecular pathways and physiological roles. Physiological Reviews. — PubMed: Labunskyy 2014
- Brigelius-Flohé R, Maiorino M (2013). Glutathione peroxidases. BBA. — PubMed: GPx family review
- Conrad M, Pratt DA (2019). The chemical basis of ferroptosis. Nature Chemical Biology. — PubMed: Ferroptosis chemistry
- Burk RF, Hill KE (2015). Regulation of selenium metabolism and transport. Annual Review of Nutrition. — PubMed: Burk Hill 2015
- Berry MJ, Banu L, Chen YY, et al. (1991). Recognition of UGA as a selenocysteine codon in type I deiodinase requires sequences in the 3' untranslated region. Nature. — PubMed: SECIS discovery
- Schomburg L, Schweizer U (2009). Hierarchical regulation of selenoprotein expression. BBA. — PubMed: Selenoprotein hierarchy
- Yant LJ et al. (2003). The selenoprotein GPx4 is essential for mouse development. FRBM. — PubMed: GPx4 embryonic lethality
- Yamashita Y, Yamashita M (2010). Selenoneine, the predominant chemical form of organic selenium in tuna blood. J Biol Chem. — PubMed: Selenoneine discovery
- Ip C, Lisk DJ (1995). Bioactivity of selenium from Brazil nut for cancer prevention. Nutr Cancer. — PubMed: Brazil nut bioactivity
- MacFarquhar JK et al. (2010). Acute selenium toxicity associated with a dietary supplement. Arch Intern Med. — PubMed: Selenium toxicity outbreak
Research Papers: Thyroid Function
- Köhrle J (2010). Selenium and the thyroid. Thyroid. — PubMed: Kohrle review
- Gartner R et al. (2002). Selenium supplementation in patients with autoimmune thyroiditis decreases TPO antibodies. JCEM. — PubMed: Gartner 2002
- Marcocci C et al. (2011). Selenium and the course of mild Graves' orbitopathy (EUGOGO). NEJM. — PubMed: EUGOGO orbitopathy trial
- Negro R et al. (2007). Selenium supplementation and postpartum thyroid status. JCEM. — PubMed: Postpartum thyroiditis
- van Zuuren EJ et al. (2013). Selenium supplementation for Hashimoto's thyroiditis (Cochrane). — PubMed: Cochrane Hashimoto's
- Wichman J et al. (2016). Selenium supplementation significantly reduces thyroid autoantibodies in chronic autoimmune thyroiditis: meta-analysis. Thyroid. — PubMed: Wichman meta-analysis
- Duntas LH, Benvenga S (2015). Selenium: an element for life. Endocrine. — PubMed: Duntas review
- Schomburg L (2017). Dietary selenium and human health. Nutrients. — PubMed: Schomburg dietary review
- Larsen PR, Davies TF (2003). Hypothyroidism and thyroiditis. Williams Textbook. — PubMed: Hypothyroidism reference
- Bianco AC, Salvatore D, Gereben B, Berry MJ, Larsen PR (2002). Biochemistry, cellular and molecular biology, and physiological roles of the iodothyronine selenodeiodinases. Endocrine Reviews. — PubMed: Deiodinase review
Research Papers: Immune Function and Viral Mutation
- Beck MA et al. (1995). Rapid genomic evolution of non-virulent Coxsackievirus B3 in selenium-deficient mice. Nature Medicine. — PubMed: Beck 1995 Nature Medicine
- Beck MA et al. (2001). Selenium deficiency increases the pathology of an influenza virus infection. FASEB Journal. — PubMed: Beck 2001 influenza
- Levander OA, Beck MA (1997). Interacting nutritional and infectious etiologies of Keshan disease. Biol Trace Elem Res. — PubMed: Keshan etiology
- Hurwitz BE et al. (2007). HIV viral load suppression with selenium supplementation. Archives of Internal Medicine. — PubMed: Hurwitz HIV trial
- Baker DH et al. (1997). Increased mortality in HIV-infected adults with low serum selenium. JAIDS. — PubMed: Baker HIV mortality
- Broome CS et al. (2004). Increased selenium intake improves immune function and poliovirus handling. AJCN. — PubMed: Broome polio trial
- Angstwurm MWA et al. (2007). Selenium in Intensive Care: high-dose selenium in severe sepsis. Crit Care Med. — PubMed: SIC trial
- Bloos F et al. (2016). Sodium selenite in severe sepsis or septic shock: SISPCT. JAMA Internal Medicine. — PubMed: SISPCT
- Hoffmann PR, Berry MJ (2008). The influence of selenium on immune responses. Mol Nutr Food Res. — PubMed: Hoffmann review
- Avery JC, Hoffmann PR (2018). Selenium, selenoproteins, and immunity. Nutrients. — PubMed: Avery Hoffmann 2018
Research Papers: Cancer Prevention (NPC, SELECT)
- Clark LC et al. (1996). Effects of selenium supplementation for cancer prevention — the NPC Trial. JAMA. — PubMed: NPC trial
- Lippman SM et al. (2009). SELECT trial of selenium and vitamin E for prostate cancer. JAMA. — PubMed: SELECT trial
- Klein EA et al. (2011). Vitamin E and prostate cancer in SELECT. JAMA. — PubMed: SELECT vitamin E
- Kristal AR et al. (2014). Baseline selenium status and SELECT prostate cancer. JNCI. — PubMed: Kristal baseline analysis
- Stranges S et al. (2007). Long-term selenium supplementation and incidence of type 2 diabetes (NPC follow-up). Annals of Internal Medicine. — PubMed: NPC diabetes signal
- Vinceti M et al. (2018). Selenium for preventing cancer (Cochrane review). — PubMed: Cochrane cancer prevention
- Hatfield DL et al. (2014). Selenium and selenocysteine: roles in cancer, health, development. Trends Biochem Sci. — PubMed: Hatfield review
- Steinbrenner H, Speckmann B, Sies H (2013). Selenium for cancer prevention: success and failures. Antioxid Redox Signal. — PubMed: Steinbrenner review
- Cai X et al. (2016). Selenium exposure and cancer risk: meta-analysis. Sci Rep. — PubMed: Cai meta-analysis
- Ip C, Hayes C, Budnick RM, Ganther HE (1991). Chemical form of selenium, critical metabolites, and cancer prevention. Cancer Res. — PubMed: Methylselenol mechanism
Research Papers: Cross-Cutting (Status, Toxicity, Form)
- Rayman MP (2012). Selenium and human health. The Lancet. — PubMed: Rayman Lancet review
- Combs GF Jr (2015). Biomarkers of selenium status. Nutrients. — PubMed: Status biomarkers
- Schomburg L, Schweizer U, Holtmann B, Bauer K, Sendtner M, Köhrle J (2003). Gene disruption discloses role of selenoprotein P in selenium delivery to target tissues. Biochem J. — PubMed: SELENOP knockout
- Combs GF Jr, Watts JC, Jackson MI, et al. (2011). Determinants of selenium status in healthy adults. Nutr J. — PubMed: Combs determinants
- Behne D, Kyriakopoulos A (2001). Mammalian selenium-containing proteins. Annu Rev Nutr. — PubMed: Selenoprotein inventory
- Bleys J, Navas-Acien A, Guallar E (2008). Serum selenium levels and all-cause, cancer, and cardiovascular mortality. Arch Intern Med. — PubMed: Bleys U-shaped mortality
- Combs GF Jr (2001). Selenium in global food systems. Br J Nutr. — PubMed: Combs global food systems
- Burk RF (2002). Selenium, an antioxidant nutrient. Nutr Clin Care. — PubMed: Burk overview
- Schomburg L (2011). Selenium, selenoproteins and the thyroid gland: interactions in health and disease. Nat Rev Endocrinol. — PubMed: Schomburg thyroid review
- Yang G, Yin S, Zhou R, et al. (1989). Studies of safe maximal daily dietary Se-intake in a seleniferous area in China. J Trace Elem Electrolytes Health Dis. — PubMed: China seleniferous toxicity
External Authoritative Resources
- Linus Pauling Institute — Selenium Micronutrient Information Center — the most comprehensive evidence-based summary of selenium biology, regularly updated
- NIH Office of Dietary Supplements — Selenium Fact Sheet for Health Professionals
- WHO — Selenium reference values and global status
- MedlinePlus — Selenium
- PubMed — All research on selenium / selenocysteine / selenoprotein
Connections
- Selenium (Main Page)
- Selenium for Antioxidant Defense
- Selenium for Cancer Prevention
- Selenium for Immune Function
- Selenium and Thyroid Function
- Selenium and Cancer Prevention — selenoprotein biology, the NPC and SELECT trials, evidence across prostate/colorectal/lung cancers, and the U-shaped dose-response curve.
- All Minerals
- Iodine
- Zinc
- Iron
- Vitamin E
- Vitamin C
- Vitamin D3
- Glutathione
- Oxidative Stress
- Immune Boosting
- Methionine
- Cysteine
- Hashimoto's Thyroiditis
- Graves' Disease
- Cancer
- Prostate Conditions
- Cardiomyopathy (Keshan)
- Organ Meats
- Tuna
- Eggs
- Garlic