Selenium for Antioxidant Defense
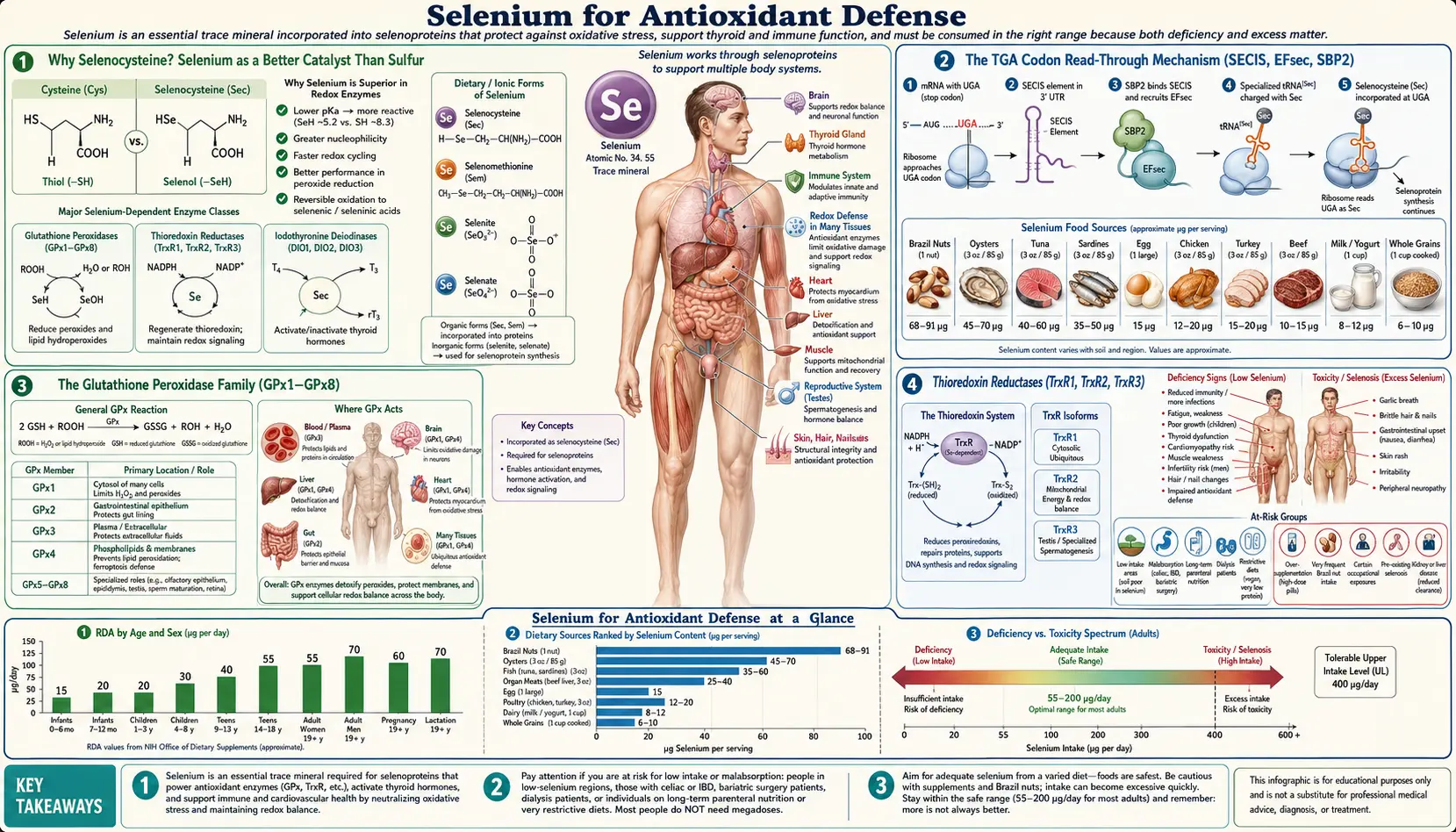
Selenium is the only trace element with its own dedicated amino acid — selenocysteine, the so-called 21st amino acid. Unlike the other 20 amino acids whose codons are universally fixed across the genetic code, selenocysteine is inserted in response to a UGA codon that ribosomes ordinarily read as STOP. A specialized 3'-untranslated SECIS hairpin and a dedicated selenocysteine-tRNA force a context-dependent read-through, allowing the cell to slot the seleno-amino acid into the catalytic site of 25 human selenoproteins. The resulting selenoenzymes — the glutathione peroxidases (GPx1-8), thioredoxin reductases (TrxR1-3), methionine sulfoxide reductase B1, and selenoprotein P — constitute the cell's most catalytically efficient redox defense. This deep dive walks through that elegant biology, the eight glutathione peroxidase isoforms (each guarding a different cellular compartment), the strange biochemistry of dietary selenoamino acids, the wide soil-driven variability of Brazil nuts, and the recently discovered marine selenoneine that concentrates in tuna mitochondria.
Table of Contents
- Why Selenocysteine? Selenium as a Better Catalyst Than Sulfur
- The TGA Codon Read-Through Mechanism (SECIS, EFsec, SBP2)
- The Glutathione Peroxidase Family (GPx1–GPx8)
- Thioredoxin Reductases (TrxR1, TrxR2, TrxR3)
- Methionine Sulfoxide Reductase B1 (MsrB1)
- Selenoprotein P: The Selenium Transport Vehicle
- Dietary Selenoamino Acids (SeMet, SeCys, MeSeCys)
- Brazil Nut Variability and Selenium Toxicity
- Selenoneine in Tuna and the Marine Mitochondrial Compartment
- Oxidative-Stress-Driven Disorders and the Selenium Connection
- Cautions and the Narrow Therapeutic Window
- Key Research Papers
- Connections
- Featured Videos
Why Selenocysteine? Selenium as a Better Catalyst Than Sulfur
Sulfur and selenium sit one row apart in column 16 of the periodic table, and their chemistry is closely related — both form bonds of comparable geometry, both can shuttle through several oxidation states, and both anchor reactive groups at protein active sites. From a distance, swapping a selenium for a sulfur looks like a minor edit. Yet across hundreds of millions of years of evolution, organisms from bacteria to mammals have invested an enormous translational apparatus (described in the next section) to make sure that one particular cysteine residue at the active site of a small set of enzymes is built as selenocysteine rather than cysteine. The reason is straightforward catalytic chemistry.
- Lower pKa – The selenol group (-SeH) has a pKa of about 5.2, compared to about 8.5 for the thiol group (-SH) of cysteine. At physiological pH (7.4), selenocysteine sits as the deprotonated, nucleophilic selenolate (-Se−), while cysteine is mostly protonated and unreactive. The nucleophilic selenolate attacks hydroperoxide substrates orders of magnitude faster than the cysteine thiolate.
- Larger atomic radius and polarizability – Selenium's larger electron cloud allows it to stabilize partial charges in the transition state of redox reactions, lowering the activation energy further.
- More reversible oxidation states – The Sec-Se(O)H selenenic acid intermediate that forms when the selenolate reduces a peroxide is more readily re-reduced (by glutathione or thioredoxin) than the analogous cysteine sulfenic acid. The enzyme can therefore turn over many catalytic cycles per second without being trapped in an over-oxidized dead-end state — the Sec-SeO2H (seleninic acid) and Sec-SeO3H (selenonic acid) over-oxidized forms are far rarer than the Cys-SO2H and Cys-SO3H states that permanently inactivate purely-cysteine peroxidases under oxidative stress.
The net effect: selenocysteine-containing enzymes catalyze peroxide reduction roughly 100 to 1000 times faster than cysteine-only homologs, and they are far more resistant to substrate-driven inactivation. Evolution apparently judged that catalytic advantage worth the enormous machinery cost of context-dependent UGA read-through.
The TGA Codon Read-Through Mechanism (SECIS, EFsec, SBP2)
The genetic code is conventionally described as having 61 sense codons (specifying the 20 standard amino acids) and 3 stop codons (UAA, UAG, UGA). Selenocysteine breaks that rule: UGA, which the ribosome normally reads as a translation-termination signal, is recontextually read as selenocysteine in the presence of a specific 3'-untranslated mRNA structure plus a dedicated suite of accessory proteins.
The cellular machinery required for selenocysteine insertion includes:
- tRNA[Ser]Sec – A unique tRNA that is initially charged with serine by seryl-tRNA synthetase, then chemically converted on the tRNA itself to phosphoserine (by PSTK kinase) and then to selenocysteine (by SepSecS) using selenophosphate as the selenium donor. This is the only known case where an amino acid is built directly on a tRNA rather than being attached in its finished form.
- Selenophosphate synthetase 2 (SEPHS2/SPS2) – Itself a selenoprotein, SEPHS2 generates the selenium donor molecule monoselenophosphate (H2SePO3-) from ATP and selenide. The fact that the enzyme that makes the selenium donor is itself a selenoprotein creates a tight feedback loop between selenium supply and selenoprotein synthesis capacity.
- SECIS element (selenocysteine insertion sequence) – A specific stem-loop RNA structure in the 3'-untranslated region of every selenoprotein mRNA. The SECIS element recruits the protein factors needed to redirect ribosomal interpretation of the upstream UGA from stop to sense.
- SBP2 (SECIS-binding protein 2) – Binds the SECIS hairpin and provides a docking surface for EFsec and the selenocysteyl-tRNA.
- EFsec (eukaryotic elongation factor specific for selenocysteine) – The dedicated GTP-binding elongation factor that delivers the selenocysteyl-tRNA to the ribosomal A-site when UGA is encountered in the context of a downstream SECIS.
The complexity of this system means that selenoprotein synthesis has a built-in pause point: when cellular selenium is scarce, the limiting factor is selenophosphate, and ribosomes that read through to a UGA without successfully recruiting a selenocysteyl-tRNA simply terminate, producing a truncated, non-functional protein that is then degraded. This is why selenium deficiency does not just reduce selenoenzyme activity proportionally — it produces a hierarchy of selenoprotein expression in which "stress" selenoproteins like GPx1 are sacrificed first, while "housekeeping" selenoproteins like GPx4 and thioredoxin reductases are preserved as long as possible. The brain, testes, and endocrine glands sit at the top of this hierarchy and are protected last; the liver and kidney lose selenoprotein expression first during deficiency.
The Glutathione Peroxidase Family (GPx1–GPx8)
The glutathione peroxidase (GPx) family is the largest and most clinically important class of selenoproteins. Eight human GPx isoforms are recognized, of which five (GPx1, GPx2, GPx3, GPx4, GPx6) contain selenocysteine at their active site and three (GPx5, GPx7, GPx8) contain ordinary cysteine and are therefore much less efficient peroxidases. Each isoform occupies a specific cellular or tissue compartment, and the deletion of any one of them in mice produces a distinct disease phenotype.
- GPx1 (cytosolic, ubiquitous) – The most abundant selenoprotein in most cells. GPx1 scavenges H2O2 in the cytoplasm using reduced glutathione as the electron donor. GPx1-knockout mice are viable but show increased susceptibility to oxidative stress, ischemia-reperfusion injury, and viral-induced cardiomyopathy.
- GPx2 (gastrointestinal epithelium) – Expressed in the intestinal lining, GPx2 defends against dietary pro-oxidants, microbial metabolites, and inflammation-driven peroxide accumulation. Combined GPx1/GPx2 double-knockout mice develop spontaneous ileocolitis.
- GPx3 (extracellular, plasma) – The major extracellular selenoenzyme, GPx3 is secreted primarily by the kidney and the thyroid follicular lumen. In the thyroid, GPx3 quenches excess H2O2 generated by DUOX1/DUOX2 during thyroid hormone synthesis — the same mechanism described in detail on the Selenium and Thyroid Function page. GPx3 promoter methylation silences its expression in many epithelial cancers.
- GPx4 (phospholipid hydroperoxide GPx) – The structurally distinct GPx that can directly reduce phospholipid hydroperoxides embedded in membranes, cholesterol hydroperoxides, and thymine hydroperoxides in DNA. GPx4 is now recognized as the master regulator of ferroptosis, an iron-dependent, lipid-peroxidation-driven form of regulated cell death distinct from apoptosis and necroptosis. GPx4 knockout is embryonically lethal at gastrulation, the most severe phenotype of any selenoprotein deletion. In adults, GPx4-deficient neurons die by ferroptosis, and GPx4-deficient sperm have severely impaired motility because the mitochondrial capsule of the sperm midpiece is built from cross-linked GPx4 polymers.
- GPx5 (epididymis, cysteine-only) – Expressed in the male reproductive tract and protects spermatozoa during epididymal maturation.
- GPx6 (olfactory epithelium and embryo) – Selenocysteine-containing in humans but cysteine-only in rodents; its function in olfactory neurogenesis is incompletely characterized.
- GPx7 and GPx8 (endoplasmic reticulum, cysteine-only) – Coupled to the protein-disulfide isomerase machinery, these isoforms reduce the H2O2 generated as a by-product of disulfide-bond formation during nascent-protein folding in the ER lumen.
The clinical takeaway is that selenium deficiency is not a single-enzyme deficit but a compartment-by-compartment failure of the redox-buffering network — cytosol (GPx1), gut lumen (GPx2), plasma and thyroid follicle (GPx3), membranes (GPx4), reproductive tract (GPx5), and the ER (GPx7/8). The phenotypic consequences depend on which compartment fails first under a given stressor.
Thioredoxin Reductases (TrxR1, TrxR2, TrxR3)
The thioredoxin (Trx) system runs in parallel to the glutathione (GSH) system and is in many cellular contexts the more important of the two redox-buffering networks. Trx is a small redox protein that uses a vicinal Cys-Gly-Pro-Cys active-site motif to reduce disulfides on substrate proteins. Oxidized Trx is then itself re-reduced by the selenoenzyme thioredoxin reductase (TrxR), using NADPH as the ultimate electron donor.
- TrxR1 (cytosolic and nuclear) – The principal cytoplasmic isoform. TrxR1 reduces cytosolic Trx1, which in turn reduces peroxiredoxins (Prx1, Prx2, Prx4), the redox-regulated transcription factors NF-kB, AP-1, p53, and the iron-binding protein ferritin. TrxR1 also reduces oxidized ribonucleotide reductase, the enzyme that builds the deoxyribonucleotide pool required for DNA synthesis — making selenium status a rate-limiting factor for cell proliferation and DNA repair.
- TrxR2 (mitochondrial) – Reduces mitochondrial Trx2 and the mitochondrial peroxiredoxin Prx3. TrxR2 is the chief defense against the constant flux of superoxide and hydrogen peroxide generated by the electron transport chain. TrxR2 knockout is embryonically lethal at midgestation with severe cardiac and hematopoietic defects.
- TrxR3 / TGR (testis, cytosolic and ER) – A dual-domain enzyme combining thioredoxin reductase and glutaredoxin activities. Highly expressed in spermatogenic cells, where it participates in the formation of the disulfide bonds that crosslink sperm structural proteins.
An interesting therapeutic complication: TrxR1 is overexpressed in many tumors, and a number of anticancer compounds (auranofin, motexafin gadolinium, certain platinum complexes, ethaselen) work in part by inhibiting TrxR1, collapsing the redox buffer that cancer cells rely on to survive their elevated baseline oxidative stress. This is part of the rationale for the apoptosis-selective effects of high-dose selenium metabolites on cancer cells described on the Selenium and Cancer Prevention page.
Methionine Sulfoxide Reductase B1 (MsrB1)
Methionine residues in proteins are particularly susceptible to oxidation by reactive oxygen species. The oxidation product, methionine sulfoxide, has two possible diastereomers (S-MetO and R-MetO), and a different enzyme is required to reduce each. MsrA reduces S-MetO; the family of MsrB enzymes reduces R-MetO. Of the three MsrB isoforms in humans (MsrB1, MsrB2, MsrB3), only MsrB1 is a selenoprotein; MsrB2 and MsrB3 use ordinary cysteine and are correspondingly less active.
The functional significance of methionine sulfoxide reductases is twofold. First, they directly repair oxidized methionine residues, restoring protein function. Second, surface-exposed methionines on proteins act as a sacrificial antioxidant buffer — they can be oxidized and re-reduced repeatedly, absorbing oxidative hits that would otherwise damage more critical residues. The methionine-MsrB1 cycle is therefore one of the few protein-level antioxidant systems that is truly reversible at biological timescales. Loss of MsrB1 activity in selenium deficiency contributes to the accumulation of oxidized, dysfunctional proteins and accelerates the formation of the protein aggregates implicated in neurodegenerative disease.
Selenoprotein P: The Selenium Transport Vehicle
Selenoprotein P (SELENOP) is the single most unusual selenoprotein in the human proteome: it contains up to ten selenocysteine residues per molecule (compared to one Sec per molecule for every other selenoprotein), and its primary function is to deliver selenium from the liver to peripheral tissues. SELENOP is produced almost entirely by hepatocytes and secreted into the bloodstream; its plasma concentration is the most sensitive single biomarker of long-term whole-body selenium status, more reliable than plasma GPx3 activity or serum selenium itself.
Peripheral uptake of SELENOP is receptor-mediated and tissue-specific:
- Apolipoprotein E receptor 2 (ApoER2 / LRP8) – The major SELENOP receptor in the brain (across the blood-brain barrier endothelium and at the choroid plexus) and the testes. ApoER2 internalizes SELENOP, the lysosomal protease cathepsin B then degrades it into selenocysteine residues, and the released selenocysteine is processed by selenocysteine lyase into selenide for new selenoprotein synthesis. ApoER2-knockout mice show severe selenium deficiency in brain and testis even when whole-body selenium status is normal — selenium gets to the periphery but cannot be delivered into the protected compartments.
- Megalin (LRP2) – The proximal-tubule receptor that reabsorbs SELENOP filtered at the glomerulus. Without megalin-mediated reabsorption, all SELENOP would be lost in urine.
SELENOP also has direct antioxidant activity in the extracellular space — its C-terminal selenocysteines can reduce phospholipid hydroperoxides in lipoproteins and on endothelial membranes, providing a first line of defense against oxidative damage in the vascular compartment.
Dietary Selenoamino Acids (SeMet, SeCys, MeSeCys)
Selenium does not enter the body as the free element — it enters as one of three principal selenoamino acids, each with distinct metabolism and biological fate.
- Selenomethionine (SeMet) – The principal selenium form in plant foods, fortified yeast, Brazil nuts, fish, and meat. Plants and yeast cannot distinguish selenate or selenite from sulfate or sulfite at the level of sulfur metabolism, so they incorporate dietary selenium non-specifically in place of sulfur, producing SeMet wherever methionine would otherwise sit. Once absorbed, SeMet is similarly incorporated non-specifically into body proteins wherever methionyl-tRNA is loaded — the protein-bound SeMet pool represents the body's main selenium reserve. Direct delivery of SeMet selenium into selenoprotein synthesis requires first the catabolic release of selenide from SeMet via the transsulfuration / methionine cycle. SeMet has the highest bioavailability of any dietary selenium form (about 90%) and produces the most gradual, durable rise in tissue selenium content.
- Selenocysteine (SeCys) – Released by digestive proteolysis of dietary selenoproteins (mostly from animal sources). SeCys cannot be re-incorporated into proteins by the standard translational machinery (which only handles the co-translational selenocysteyl-tRNA pathway described above), so it is funneled directly into selenide by selenocysteine lyase. Selenide is then used to make monoselenophosphate, which is then used to build new selenocysteyl-tRNA, which is then co-translationally inserted into newly synthesized selenoproteins. SeCys-derived selenium therefore reaches the selenoprotein pool faster than SeMet-derived selenium and is less prone to non-specific accumulation in the methionine pool.
- Methylselenocysteine (MeSeCys, Se-methylselenocysteine) – The principal selenium form in members of the Allium family (garlic, onions, leeks, ramps) and the Brassica family (broccoli, cauliflower, cabbage) when these plants are grown on selenium-enriched soil or selenium-fertilized. MeSeCys bypasses the methionine pool entirely; it is cleaved directly by β-lyase enzymes to release methylselenol (CH3SeH), the active anticancer metabolite. Methylselenol is the species that selectively induces apoptosis in transformed cells, inhibits angiogenesis, and modulates the tumor microenvironment, as discussed on the Selenium and Cancer Prevention page. Selenium-enriched broccoli and selenium-enriched garlic have been investigated specifically because they deliver MeSeCys without the side-effects of supraphysiologic selenium loading.
Brazil Nut Variability and Selenium Toxicity
Brazil nuts (Bertholletia excelsa) are widely cited as the richest dietary source of selenium — popular reference numbers cite 68 to 91 micrograms of selenium per nut. That is roughly the entire RDA (55 mcg/day) in a single nut. The reality, however, is much more variable than the textbook numbers suggest, and that variability has caused well-documented cases of selenosis in selenium-curious consumers.
- Soil-driven range – Selenium content of Brazil nuts ranges from approximately 5 to 100+ mcg per nut, driven almost entirely by the selenium concentration of the soils where the tree was grown. Nuts harvested from selenium-rich soils in central Brazil (the Amazon and Cerrado regions) routinely exceed 90 mcg/nut; nuts from selenium-depleted soils in northern South America may contain less than 10 mcg/nut. A single bag in a supermarket may contain nuts from multiple harvests and multiple regions.
- Cumulative toxicity in habitual consumers – Consumers who hear that "Brazil nuts are great for selenium" and eat 6 to 10 per day can easily exceed the 400-mcg/day tolerable upper intake (UL) within a single serving if the nuts are from a selenium-rich harvest. Documented case reports describe garlic-odor breath, brittle nails, hair loss, fatigue, peripheral neuropathy, and nausea in patients eating multiple Brazil nuts per day for months.
- Practical guidance – One to two Brazil nuts per day is generally sufficient for selenium repletion. Eating more than that on a sustained basis is not safer; it is the only reasonably common way for an ordinary American adult to develop true selenosis.
Selenoneine in Tuna and the Marine Mitochondrial Compartment
Most discussion of dietary selenium stops at the three classical selenoamino acids above. A more recent discovery, with implications for understanding mercury detoxification in marine ecosystems, is selenoneine — a selenium-containing analog of ergothioneine that is the dominant chemical form of selenium in the red muscle of tuna and other large pelagic predators.
- Structural relationship to ergothioneine – Ergothioneine is a sulfur-containing histidine derivative (2-thiohistidine betaine) that concentrates in mitochondria via the OCTN1 / SLC22A4 transporter and serves as a tissue-specific antioxidant in the high-oxidative-stress compartments of brain, eye, liver, and red blood cells. Selenoneine is the selenium analog — the 2-selenohistidine betaine — and shares the same OCTN1-mediated mitochondrial concentration mechanism.
- Concentration in marine predators – Tuna red muscle contains selenoneine at concentrations of 0.5 to 8 mg/kg, corresponding to roughly 50 to 90 percent of total selenium in some species (bluefin and skipjack tuna). Lower-trophic-level fish (sardines, herring) contain less, and freshwater fish and terrestrial meat contain almost none.
- Mercury detoxification hypothesis – Selenoneine forms a very stable selenoneine-mercury complex (an HgSe-protein-bound species), and this is one of the principal proposed mechanisms by which marine fish detoxify dietary methylmercury. The "selenium-to-mercury molar ratio" concept that has emerged in fish-toxicology guidelines is driven in part by selenoneine biology.
- Implications for human consumers – Selenoneine is bioavailable in humans and produces a distinct, persistent rise in red-blood-cell selenium that does not respond to standard SeMet or selenite supplementation. Regular consumers of tuna and other large predatory fish accumulate significant selenoneine in their RBCs, potentially providing an additional layer of mitochondrial antioxidant defense not seen in non-fish-eating populations.
Oxidative-Stress-Driven Disorders and the Selenium Connection
Selenium deficiency does not produce a single textbook disease in the way that vitamin C deficiency produces scurvy or vitamin D deficiency produces rickets. Instead, it acts as a permissive factor that worsens the cellular consequences of unrelated stressors. Several clinical syndromes illustrate the pattern.
- Keshan disease (selenium-deficient cardiomyopathy) – An endemic dilated cardiomyopathy first described in selenium-deficient regions of China. Selenium deficiency alone is necessary but apparently not sufficient — the trigger appears to be a normally benign Coxsackievirus B3 infection that mutates to a virulent cardiotoxic form in selenium-deficient hosts. This Keshan story is the canonical example of selenium's role in viral mutation prevention and is discussed in detail on the Selenium and Immune Function page.
- Kashin-Beck disease – An osteoarthropathy with cartilage necrosis prevalent in selenium- and iodine-deficient regions of China, Tibet, and Siberia. Cartilage chondrocytes are particularly dependent on GPx and TrxR activity for redox-driven extracellular matrix maintenance.
- Ferroptosis-driven neurodegeneration – GPx4 is the master regulator of ferroptosis, and acute GPx4 inactivation in adult neurons triggers iron-dependent lipid peroxidation that drives a distinct form of programmed cell death. The relevance to Parkinson's disease, ALS, Huntington's disease, and certain hemorrhagic-stroke contexts is an active area of research.
- Sperm motility loss – GPx4-deficient sperm have severely defective mitochondrial capsules in the midpiece and cannot generate the ATP needed for sustained flagellar beat. Selenium deficiency is a recognized contributor to male infertility worldwide.
- Hashimoto's and Graves' orbitopathy – Covered in detail on the Thyroid Function page.
Cautions and the Narrow Therapeutic Window
- Tolerable Upper Intake Level (UL) is only 400 mcg/day – among the lowest ULs of any essential nutrient. The therapeutic window between the 55-mcg/day RDA and the 400-mcg/day UL is roughly 7-fold, narrower than for any other essential trace element.
- Acute selenosis – Garlic-odor breath, metallic taste, nausea, GI distress, and CNS irritability after single high-dose ingestion. Severe cases require gastric decontamination and hospitalization.
- Chronic selenosis – Brittle nails with characteristic Mees-line transverse white bands, hair loss (often patchy), skin rash, peripheral neuropathy, and fatigue with chronic intake above the UL. The classic clinical setting is a patient consuming 5-10 Brazil nuts per day for months.
- NPC trial type-2 diabetes signal – The long-term follow-up of the Nutritional Prevention of Cancer trial found a small but statistically significant increase in incident type 2 diabetes in the 200-mcg/day selenium arm among participants who were selenium-replete at baseline. This was the first major safety signal that "more selenium" is not better in adequate populations.
- SELECT trial null result and prostate-cancer signal – The much larger SELECT trial found no cancer prevention benefit from 200 mcg/day selenomethionine in a selenium-replete population, and a non-significant increase in high-grade prostate cancer in the selenium-plus-vitamin-E arm prompted early termination. The take-away: supplementing selenium above adequate levels does not help and may hurt.
- Drug interactions – May enhance the effect of anticoagulants. Should be separated from levothyroxine dosing by several hours to avoid binding interference. Consider conservative dosing in advanced chronic kidney disease (impaired SELENOP clearance).
- Pregnancy – The RDA increases modestly to 60 mcg/day in pregnancy and 70 mcg/day in lactation, but supraphysiologic supplementation in pregnancy has not been shown safe; do not exceed the 400-mcg/day UL.
This content is provided for informational purposes only and does not constitute medical advice. Consult a qualified healthcare provider before starting selenium supplementation, especially if you eat Brazil nuts regularly or have thyroid, kidney, or cardiovascular disease.
Key Research Papers
- Rayman MP (2012). Selenium and human health. The Lancet. — DOI: 10.1016/S0140-6736(11)61452-9
- Labunskyy VM, Hatfield DL, Gladyshev VN (2014). Selenoproteins: molecular pathways and physiological roles. Physiological Reviews. — DOI: 10.1152/physrev.00039.2013
- Brigelius-Flohé R, Maiorino M (2013). Glutathione peroxidases. Biochim Biophys Acta. — DOI: 10.1016/j.bbagen.2012.11.020
- Conrad M, Pratt DA (2019). The chemical basis of ferroptosis. Nature Chemical Biology. — DOI: 10.1038/s41589-019-0408-1
- Yant LJ et al. (2003). The selenoprotein GPx4 is essential for mouse development and protects from radiation and oxidative damage insults. Free Radical Biology & Medicine. — DOI: 10.1016/S0891-5849(03)00114-2
- Burk RF, Hill KE (2015). Regulation of selenium metabolism and transport. Annual Review of Nutrition. — DOI: 10.1146/annurev-nutr-071714-034250
- Berry MJ, Banu L, Chen YY, et al. (1991). Recognition of UGA as a selenocysteine codon in type I deiodinase requires sequences in the 3' untranslated region. Nature. — DOI: 10.1038/353273a0
- Copeland PR, Fletcher JE, Carlson BA, et al. (2000). A novel RNA binding protein, SBP2, is required for the translation of mammalian selenoprotein mRNAs. EMBO Journal. — DOI: 10.1093/emboj/19.2.306
- Schomburg L, Schweizer U (2009). Hierarchical regulation of selenoprotein expression and sex-specific effects of selenium. Biochim Biophys Acta. — DOI: 10.1016/j.bbagen.2009.03.029
- Burk RF, Hill KE (2009). Selenoprotein P — expression, functions, and roles in mammals. Biochim Biophys Acta. — DOI: 10.1016/j.bbagen.2009.03.026
- Yamashita Y, Yamashita M (2010). Identification of a novel selenium-containing compound, selenoneine, as the predominant chemical form of organic selenium in the blood of bluefin tuna. J Biol Chem. — DOI: 10.1074/jbc.C110.106377
- Ip C, Lisk DJ (1995). Bioactivity of selenium from Brazil nut for cancer prevention and selenoenzyme maintenance. Nutrition and Cancer. — DOI: 10.1080/01635589509514392
- MacFarquhar JK et al. (2010). Acute selenium toxicity associated with a dietary supplement. Archives of Internal Medicine. — DOI: 10.1001/archinternmed.2009.495
- Stranges S et al. (2007). Effects of long-term selenium supplementation on the incidence of type 2 diabetes: a randomized trial. Annals of Internal Medicine. — DOI: 10.7326/0003-4819-147-4-200708210-00175
PubMed Topic Searches
- PubMed: Selenocysteine UGA SECIS readthrough
- PubMed: GPx4 and ferroptosis
- PubMed: Thioredoxin reductase selenoprotein
- PubMed: Selenoprotein P and ApoER2 brain transport
- PubMed: Methylselenocysteine chemoprevention
- PubMed: Selenoneine tuna mercury
- PubMed: Brazil nut selenosis
Connections
- Selenium Benefits Hub
- Selenium (Main Page)
- Selenium for Cancer Prevention
- Selenium for Immune Function
- Selenium and Thyroid Function
- Selenium and Cancer Prevention — selenoprotein biology, the NPC and SELECT trials, evidence across prostate/colorectal/lung cancers, and the U-shaped dose-response curve.
- Glutathione
- Oxidative Stress
- Methionine
- Cysteine
- Zinc
- Iodine
- Vitamin E
- Vitamin C
- Garlic
- Broccoli Sprouts
- Organ Meats
- Tuna
- Eggs
- Mercury