Iron and Athletic Performance
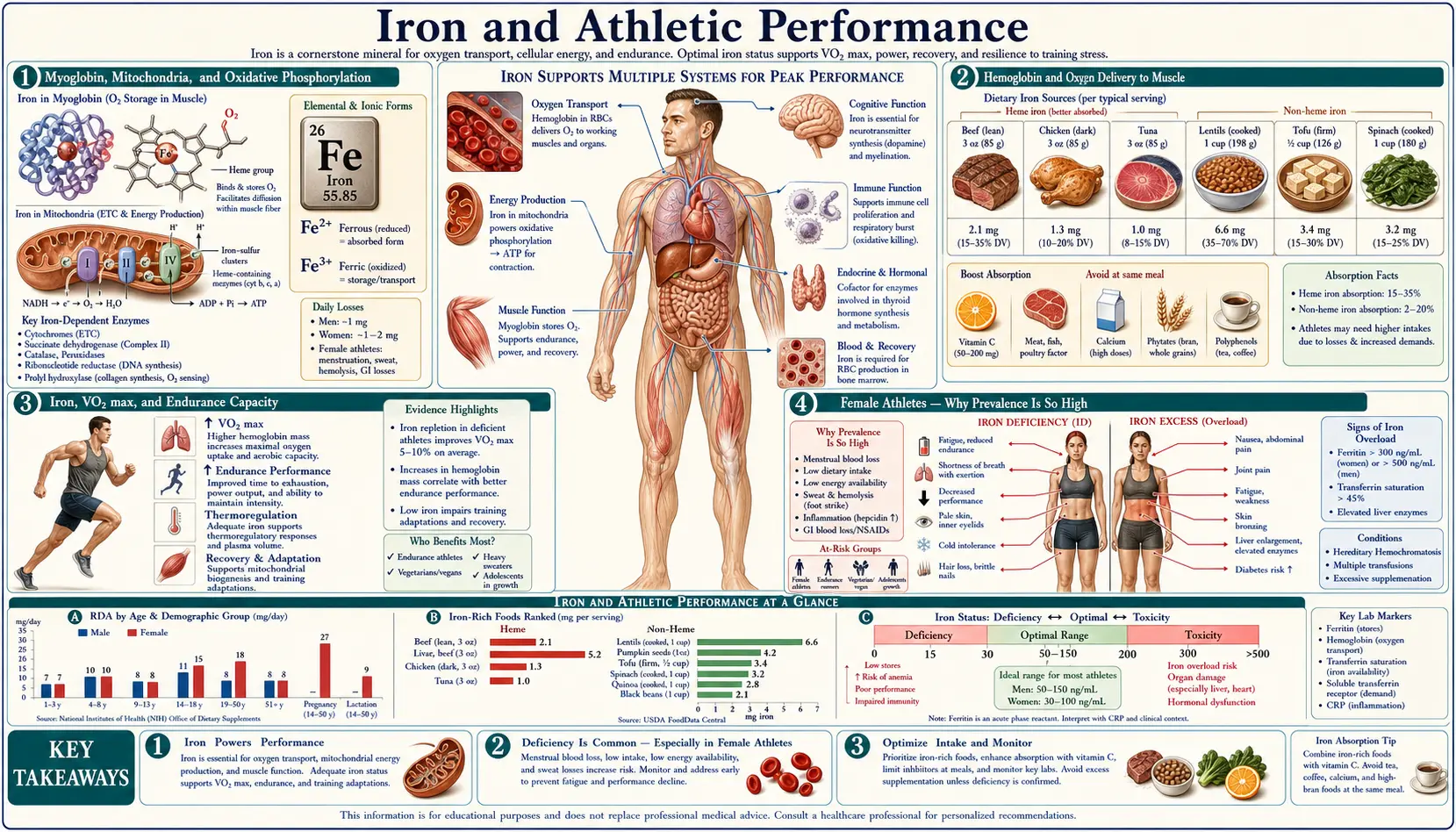
Iron is the single nutrient with the largest documented impact on endurance athletic performance. Hemoglobin carries oxygen from lung to muscle, myoglobin buffers oxygen within the working muscle, and the iron-sulfur clusters and heme cytochromes of the mitochondrial electron transport chain extract energy from that oxygen. Iron deficiency at any of these levels reduces VO2max, time to exhaustion, and submaximal endurance performance — effects that appear at the iron-deficient-erythropoiesis stage, before hemoglobin drops below the WHO anemia cutoff. Female endurance athletes are the highest-risk group, with iron-deficiency prevalence estimates of 20-50% in collegiate distance runners, rowers, and triathletes. The Brutsaert and Beard intervention trials demonstrated VO2max and time-trial improvements with iron repletion in non-anemic iron-deficient women, and the Peeling-Stoffel work on the hepcidin-IL-6 axis explained why endurance training itself transiently suppresses iron absorption in the post-exercise window. This deep-dive maps the molecular biology onto practical training and supplementation strategy.
Table of Contents
- Myoglobin, Mitochondria, and Oxidative Phosphorylation
- Hemoglobin and Oxygen Delivery to Muscle
- Iron, VO2max, and Endurance Capacity
- Female Athletes — Why Prevalence Is So High
- Beard and Brutsaert Intervention Trials
- The Hepcidin-IL-6 Inflammatory Feedback in Exercise
- "Training-Induced Anemia" vs True Iron Deficiency
- Foot-Strike Hemolysis and GI Losses
- Screening, Targets, and Practical Supplementation
- Cautions (Overload, Doping Rules, Hemochromatosis)
- Key Research Papers
- Connections
- Featured Videos
Myoglobin, Mitochondria, and Oxidative Phosphorylation
Skeletal and cardiac muscle contain approximately 10% of the body's total iron, the majority of which is locked inside myoglobin and the mitochondrial electron transport chain. Myoglobin is the muscle-specific cousin of hemoglobin, a monomeric heme protein with a higher oxygen affinity than hemoglobin, which gives it the role of a local oxygen buffer — it accepts oxygen from hemoglobin in the muscle capillary bed and releases it intracellularly to the mitochondria as oxygen demand spikes during contraction. Muscle fibers with high oxidative capacity (slow-twitch type I, fast-twitch oxidative type IIa) have high myoglobin content, which is why those fibers appear red and dark. The fast-twitch glycolytic type IIb fibers have low myoglobin and appear pale.
The mitochondrial electron transport chain depends on iron at two distinct levels. Heme-containing cytochromes (b, c1, c, a, and a3) shuttle single electrons between Complexes III and IV, with Complex IV (cytochrome c oxidase) catalyzing the final four-electron reduction of O2 to water. Iron-sulfur clusters in Complexes I, II, and III also carry electrons one at a time as the gradient is built up across the inner mitochondrial membrane. Aconitase, the citric acid cycle enzyme that interconverts citrate and isocitrate, requires a [4Fe-4S] cluster for catalysis.
The practical consequence: iron deficiency degrades muscle oxidative capacity at the enzymatic level before hemoglobin drops. Skeletal muscle biopsies in iron-deficient rats show reduced cytochrome c oxidase activity, reduced ATP production, and reduced endurance time on treadmill running — all of which can be partially restored within days of starting iron repletion, before hemoglobin or hematocrit normalize. This separation between "central" (hemoglobin-mediated oxygen delivery) and "peripheral" (mitochondrial-mediated oxygen utilization) iron effects is why iron-deficient-but-non-anemic athletes can still see meaningful performance gains from iron repletion.
Hemoglobin and Oxygen Delivery to Muscle
Approximately two-thirds of total body iron resides in circulating hemoglobin, which carries oxygen from the alveolar capillaries to working tissues. The maximum rate of aerobic energy production is directly limited by the rate at which oxygen can be delivered to the mitochondria, and this delivery is the product of cardiac output, hemoglobin concentration, and arterial oxygen saturation (the Fick principle: VO2 = cardiac output × arteriovenous oxygen difference).
Each gram of hemoglobin can carry approximately 1.34 mL of oxygen when fully saturated. An adult male with 15 g/dL hemoglobin and 5 L of blood thus has a total circulating oxygen-carrying capacity of about 1 L; an iron-deficient woman with 11 g/dL hemoglobin and 4 L of blood has about 590 mL. The difference is a roughly 40% reduction in maximal oxygen-carrying capacity — equivalent in performance terms to losing roughly the oxygen advantage that altitude-acclimated elite distance runners spend years to acquire. The system partially compensates with higher cardiac output (resting tachycardia and exercise tachycardia), but this compensation has limits and itself has metabolic costs (the heart is using ATP to push more blood per beat).
Within the working muscle, oxygen extraction from arterial blood is limited by the partial-pressure gradient between capillary and mitochondrion, and by myoglobin's ability to buffer oxygen delivery to the mitochondrion during the muscle contraction phase. Iron deficiency reduces this delivery capacity at the systemic level (less hemoglobin), the local level (less myoglobin), and the consumption level (less cytochrome c oxidase).
Iron, VO2max, and Endurance Capacity
VO2max — the maximum rate of oxygen consumption a person can sustain during incremental exercise — is the single best laboratory predictor of endurance performance and is tightly coupled to hemoglobin concentration. Studies in iron-deficient elite endurance athletes show a linear relationship between hemoglobin and VO2max across the normal physiological range, with each 1 g/dL change in hemoglobin producing approximately a 4 mL/kg/min change in VO2max in trained athletes. The relationship is even more pronounced in submaximal endurance tasks where lactate threshold (the exercise intensity at which blood lactate rises rapidly) is iron-sensitive.
The lactate-threshold sensitivity is particularly notable because lactate threshold (rather than VO2max per se) is what determines time-to-exhaustion in long-duration endurance events. An iron-deficient runner running at 80% of her VO2max may be above her lactate threshold and accumulating fatigue quickly, where an iron-replete runner at the same percentage of VO2max may still be below her lactate threshold and able to sustain the pace much longer. This is why iron-deficient endurance athletes complain of disproportionate fatigue at submaximal training intensities even when they can still hit acceptable VO2max numbers in a laboratory test.
Pre-iron-repletion vs post-iron-repletion comparisons in non-anemic iron-deficient female endurance athletes consistently show 1-3% improvements in time-to-exhaustion at fixed submaximal intensity and 5-15% improvements in time-trial performance over middle-distance events (3-15 km running, 10-40 km cycling). In elite athletes where every fraction of a percent matters, these are large effects.
Female Athletes — Why Prevalence Is So High
Female endurance athletes are the highest-risk demographic for iron deficiency outside of pregnancy. Reported prevalence in published cohorts of collegiate and elite athletes runs 20-50% for non-anemic iron deficiency (ferritin < 30 ng/mL) and 5-15% for iron deficiency anemia. The drivers are additive:
- Menstrual blood loss: Baseline 0.5 mg/day average iron loss, higher with heavy menses. This alone puts the RDA at 18 mg/day for premenopausal women, double the male RDA.
- Increased iron loss in sweat, urine, GI mucosa: Endurance training elevates basal iron losses by an additional 0.5-1 mg/day.
- Foot-strike hemolysis: Discussed below; mechanical destruction of red cells in the plantar capillaries during distance running releases hemoglobin into plasma where free hemoglobin is captured by haptoglobin and removed via reticuloendothelial system, with iron loss.
- Exercise-induced GI bleeding: Endurance exercise causes splanchnic vasoconstriction and ischemic GI mucosal injury, with occult blood loss documented in 16-85% of marathon runners depending on intensity. Even small per-event losses add up over a training year.
- Hepcidin elevation post-exercise: Discussed in detail below; the inflammatory IL-6 response to endurance exercise transiently suppresses iron absorption in the 3-6 hour post-exercise window.
- Dietary patterns: Many endurance athletes follow restricted or partly vegetarian diets for body-composition or perceived-health reasons, reducing heme iron intake. Carbohydrate-loaded diets often displace red meat. Energy-deficient (REDS, female athlete triad) athletes have even lower iron intake.
- Frequent blood donation: Some athletes donate plasma or whole blood for income and inadvertently strip 200-250 mg iron per donation.
The cumulative result is that an elite female endurance athlete training 60-100 km per week on a partly-plant-based diet is essentially guaranteed to develop iron deficiency over the course of a season unless she actively supplements or monitors ferritin and intervenes. The clinical implication is that ferritin screening should be a routine part of preseason and mid-season physical exam in any competitive endurance program with female athletes, with proactive intervention rather than reactive treatment of frank anemia.
Beard and Brutsaert Intervention Trials
John Beard at Penn State and Tom Brutsaert at SUNY-Albany built the rigorous evidence base for iron repletion in non-anemic iron-deficient female athletes through a series of randomized placebo-controlled trials in the early 2000s. The Brutsaert 2003 trial randomized 41 iron-depleted but non-anemic untrained women to ferrous sulfate (100 mg elemental iron daily) or placebo for 6 weeks, then put them through a 6-week aerobic training program. The iron-supplemented group showed:
- Larger increase in VO2max with training (+12% vs +6% placebo)
- Greater improvement in maximal oxygen consumption per kg of fat-free mass
- Higher serum ferritin (mean 35 ng/mL vs 17 ng/mL)
- No difference in hemoglobin between groups (both groups remained non-anemic throughout)
The interpretation was that even in subjects not classically anemic, iron deficiency limited the ability to respond to endurance training stimulus — presumably by limiting mitochondrial biogenesis and the cytochrome / iron-sulfur-cluster expansion that normally accompanies endurance adaptation. The implication is that iron deficiency does not just hurt current performance, it hurts the capacity to improve through training.
The Hinton 2000 trial in untrained iron-depleted women had similarly produced gains in time-trial performance with iron repletion. The DellaValle work in collegiate rowers showed reduced time-trial performance correlation with ferritin status, with measurable improvement after repletion. The McClung et al. trial in Marine Corps recruits during 8 weeks of basic combat training showed iron supplementation produced measurable performance gains.
The aggregate effect-size picture: iron repletion in iron-deficient (ferritin < 20-25 ng/mL) female athletes typically produces 1-3% improvement in submaximal endurance, 2-5% improvement in VO2max, and 5-15% improvement in time-trial endurance over the 6-16 week timecourse needed to restore ferritin to > 30-40 ng/mL.
The Hepcidin-IL-6 Inflammatory Feedback in Exercise
Peter Peeling, Marc Sim, and colleagues at the University of Western Australia, along with Stephan Stoffel and others, mapped out a counterintuitive but clinically important loop: endurance exercise itself transiently elevates hepcidin, the master regulator of iron absorption and recycling, with the practical consequence that iron taken in the post-exercise window is poorly absorbed.
The mechanism: endurance exercise produces a measurable rise in interleukin-6 (IL-6) from working skeletal muscle, peaking 1-2 hours post-exercise. IL-6 is a potent stimulus for hepatic hepcidin synthesis, with hepcidin levels rising 3-6 hours post-exercise. Hepcidin binds to ferroportin (the only known cellular iron exporter) and induces its internalization and degradation, effectively closing the gate at both intestinal absorption (blocking dietary iron uptake) and macrophage iron recycling (blocking iron release from senescent red cell breakdown).
For an endurance athlete, this means that an iron-rich meal taken right after a hard workout will have substantially reduced iron absorption, while a meal taken before the workout or several hours after — ideally morning, before training — will be absorbed normally. The Stoffel et al. work has shown that iron-supplement timing (morning, pre-training) can roughly double daily iron absorption compared with evening, post-training timing in athletes with elevated hepcidin states.
The chronic version of this feedback contributes to the "anemia of endurance training" phenomenon: cumulative hepcidin elevation across consecutive training days can keep iron absorption suppressed enough to drive negative iron balance over weeks-to-months even in athletes consuming adequate dietary iron. This is a particular trap for athletes in heavy training blocks, racing seasons, or altitude camps.
The practical training-camp protocol: schedule oral iron in the morning with vitamin C and ideally separated from training by several hours. Consider alternate-day rather than daily dosing (a strategy with separate hepcidin-physiology support; the once-every-other-day dose lets hepcidin return to baseline between doses, improving fractional absorption per dose). For athletes with persistent low ferritin despite oral protocol compliance, intravenous ferric carboxymaltose (a single 750-1000 mg infusion) has become accepted practice in endurance sports medicine.
"Training-Induced Anemia" vs True Iron Deficiency
Endurance training produces an obligatory expansion of plasma volume, typically 200-400 mL within the first 1-2 weeks of a heavy training block. Because the red cell mass does not expand at the same rate, the hemoglobin concentration measured in g/dL drops — even though the absolute hemoglobin mass (total grams of hemoglobin in circulation) is unchanged or higher. This dilutional pseudoanemia is the basis of the "athletes have low hemoglobin" observation, and it is sometimes called "sports anemia" or "training-induced anemia."
The diagnostic challenge is that this pseudoanemia and true iron deficiency anemia look similar on a routine CBC — both show hemoglobin below population norms. They are distinguished by iron studies: a dilutional pseudoanemia has normal ferritin, normal transferrin saturation, and a normal red cell distribution width with normal MCV. A true iron deficiency shows low ferritin, low transferrin saturation, elevated RDW, and microcytosis on the smear.
This matters because the responses differ. Dilutional pseudoanemia is an adaptive sign of effective endurance training and should not be treated — iron supplementation in a non-deficient athlete will not improve performance. True iron deficiency anemia, even at the same hemoglobin number, should be treated promptly because performance gains are large and structural recovery improves.
The practical implication is that any time an endurance athlete shows up with a "low hemoglobin," the workup must include serum ferritin and transferrin saturation rather than starting iron empirically. Ferritin > 30 ng/mL with normal saturation = dilutional pseudoanemia, no treatment needed. Ferritin < 30 with low saturation = iron-deficient erythropoiesis, treat. Ferritin in the 30-50 ng/mL range with symptoms but no anemia = the gray zone where empirical short-term repletion trials are often appropriate based on clinical suspicion.
Foot-Strike Hemolysis and GI Losses
Foot-strike hemolysis is the mechanical destruction of red blood cells in the plantar capillaries during repetitive heel-strike of distance running. With each foot-strike, the capillaries in the heel and forefoot are compressed against the bones of the foot. The shear forces and brief ischemia damage red cell membranes, releasing free hemoglobin into the plasma where it is bound by haptoglobin and cleared by the reticuloendothelial system. A portion of the released iron is lost in urine as hemoglobinuria when haptoglobin is saturated.
The phenomenon is well-documented in marathon runners, with measurable haptoglobin depletion, free hemoglobin elevation, and occasionally visible hemoglobinuria immediately post-race. The same mechanism operates in other repetitive-impact sports including triathlon (run leg), dance, and certain military training contexts. Cycling and swimming, with low or no impact loading on the feet, do not produce foot-strike hemolysis.
Practical mitigation: cushioned shoes, soft surfaces, midfoot rather than heel strike, and lower body mass all reduce per-stride loading and thus per-race iron loss. The accumulated effect across a marathon training year is meaningful but not overwhelming — foot-strike alone does not produce iron deficiency in a runner with adequate iron intake, but it adds to other losses and tips the balance in already-marginal athletes.
Exercise-induced GI blood loss is the second mechanical loss mechanism and is often underappreciated. Splanchnic vasoconstriction during heavy endurance exercise produces ischemic mucosal injury in the gastric, small-intestinal, and colonic walls. Occult fecal blood is detectable in 16-85% of marathon runners post-race depending on intensity, hydration status, NSAID use, and individual susceptibility. Per-event losses are small, but cumulative losses over a competitive season can be significant. NSAID use, in particular, both directly damages gastric mucosa and inhibits cyclooxygenase-mediated prostaglandin protection of gastric mucosa — the routine pre-race ibuprofen of many endurance athletes is a meaningful contributor.
Screening, Targets, and Practical Supplementation
The sports-medicine consensus that has emerged over the last decade:
- Annual screening minimum: Ferritin, hemoglobin, hematocrit, MCV, RDW at preseason physical for all endurance athletes, especially female athletes.
- Mid-season recheck: For athletes with marginal baseline ferritin or symptomatic fatigue / performance decrement.
- Iron-deficient threshold (athletes): Ferritin < 35 ng/mL flags repletion candidate. Some sports-medicine programs use < 50.
- Iron-deficient anemia threshold: Hemoglobin < 12 g/dL (women) or < 13 g/dL (men) with iron-deficient iron studies — treat aggressively.
- Target after repletion: Ferritin > 50-100 ng/mL for endurance athletes, depending on training load.
The supplementation protocol that has emerged as both effective and tolerable:
- First-line: Ferrous sulfate 325 mg (65 mg elemental iron) every other day in the morning with 250 mg vitamin C and away from coffee, tea, calcium, and dairy. The alternate-day dosing improves fractional absorption (because hepcidin returns to baseline between doses) and reduces GI side effects compared with daily dosing.
- Ferrous bisglycinate alternative: 25-50 mg elemental every day or every other day for athletes who do not tolerate ferrous sulfate. Better tolerated, slightly lower mg-for-mg absorption.
- Timing relative to training: Take oral iron in the morning, at least 4-6 hours before a hard workout and at least 6-8 hours after, to avoid the post-exercise hepcidin window when absorption is suppressed.
- Intravenous ferric carboxymaltose (FCM): 750-1000 mg single dose for athletes with persistent low ferritin despite compliant oral protocol, GI intolerance, or short timeline before competition. Reasonably well-tolerated. Reserved for cases where oral fails.
- Reassessment timeline: Recheck ferritin at 8-12 weeks. Expect ferritin to rise by ~25-50 ng/mL per quarter on therapy.
- Dietary support: Heme iron sources (red meat, poultry, fish) at 3-5 meals per week. See Heme vs Non-Heme Iron.
Cautions (Overload, Doping Rules, Hemochromatosis)
- Iron overload risk: Indefinite high-dose iron supplementation in non-deficient athletes is risky. Excess iron drives Fenton-chemistry oxidative stress, accumulates in liver and heart, and in HFE-mutation carriers (1 in 200-400 of Northern European ancestry are C282Y homozygotes) drives hemochromatosis. Supplement only with documented deficiency. Stop once ferritin is in the target range. Do not maintain supplementation indefinitely.
- Pre-supplementation HFE screening: Reasonable for athletes of Northern European ancestry being started on chronic iron supplementation. C282Y/C282Y homozygotes should not be supplemented unless deficiency is documented and even then with caution and follow-up monitoring.
- WADA doping considerations: Oral and intravenous iron are not WADA-banned and have no doping classification. Erythropoietin (EPO) and EPO-mimetic peptides ARE banned. Hemoglobin and reticulocyte tracking in the Athlete Biological Passport program will flag suspicious rapid hemoglobin increases — documented iron-deficiency repletion provides a legitimate explanation but should be recorded in the athlete's medical record. Blood transfusion (autologous or homologous) is WADA-banned. There is no doping issue with normal therapeutic iron use.
- Athlete iron deficiency and disordered eating: A significant fraction of iron-deficient female endurance athletes have concurrent low energy availability (RED-S, the female athlete triad). Iron is only part of the problem. Repletion of iron without addressing energy availability is incomplete and often ineffective — the underlying restrictive eating prevents iron repletion through both inadequate intake and elevated baseline hepcidin from the relative inflammation/stress state. Comprehensive sports-medicine workup including nutrition assessment is appropriate.
- "Just push iron higher" antipattern: Once ferritin is in the 50-100 ng/mL range and performance is adequate, more iron is not better. Continuing high-dose oral iron pushes ferritin into the iron-overload range without performance benefit and with cumulative oxidative-stress and HFE-unmasking risk.
- Copper-ceruloplasmin counterpoint: See the Morley Robbins discussion on the Cognitive Performance page. Some practitioners argue that iron supplementation without copper repletion drives unbound iron deposition and underlying ceruloplasmin dysfunction. Worth checking ceruloplasmin in iron-supplementation-resistant cases.
Key Research Papers
- Brutsaert TD, Hernandez-Cordero S, Rivera J, Viola T, Hughes G, Haas JD (2003). Iron supplementation improves progressive fatigue resistance during dynamic knee extensor exercise in iron-depleted, nonanemic women. American Journal of Clinical Nutrition. — PubMed
- Hinton PS, Giordano C, Brownlie T, Haas JD (2000). Iron supplementation improves endurance after training in iron-depleted, nonanemic women. Journal of Applied Physiology. — PubMed
- Beard JL, Tobin B (2000). Iron status and exercise. American Journal of Clinical Nutrition. — PubMed
- Peeling P, Dawson B, Goodman C, Landers G, Trinder D (2008). Athletic induced iron deficiency: new insights into the role of inflammation, cytokines and hormones. European Journal of Applied Physiology. — PubMed
- Sim M, Garvican-Lewis LA, Cox GR, Govus A, McKay AKA et al. (2019). Iron considerations for the athlete: a narrative review. European Journal of Applied Physiology. — PubMed
- Stoffel NU, Cercamondi CI, Brittenham G, Zeder C, Geurts-Moespot AJ et al. (2017). Iron absorption from oral iron supplements given on consecutive versus alternate days and as single morning doses versus twice-daily split dosing in iron-depleted women. Lancet Haematology. — PubMed
- DellaValle DM, Haas JD (2011). Impact of iron depletion without anemia on performance in trained endurance athletes at the beginning of a training season. International Journal of Sport Nutrition and Exercise Metabolism. — PubMed
- McClung JP, Karl JP, Cable SJ, Williams KW, Nindl BC, Young AJ, Lieberman HR (2009). Randomized, double-blind, placebo-controlled trial of iron supplementation in female soldiers during military training. American Journal of Clinical Nutrition. — PubMed
- Telford RD, Sly GJ, Hahn AG, Cunningham RB, Bryant C, Smith JA (2003). Footstrike is the major cause of hemolysis during running. Journal of Applied Physiology. — PubMed
- Nemeth E, Rivera S, Gabayan V, Keller C, Taudorf S, Pedersen BK, Ganz T (2004). IL-6 mediates hypoferremia of inflammation by inducing the synthesis of the iron regulatory hormone hepcidin. Journal of Clinical Investigation. — PubMed
- Burden RJ, Pollock N, Whyte GP, Richards T, Moore B, Busbridge M et al. (2015). Effect of intravenous iron on aerobic capacity and iron metabolism in elite athletes. Medicine and Science in Sports and Exercise. — PubMed
- Pasricha SR, Low M, Thompson J, Farrell A, De-Regil LM (2014). Iron supplementation benefits physical performance in women of reproductive age: a systematic review and meta-analysis. Journal of Nutrition. — PubMed
PubMed Topic Searches
- PubMed: Iron deficiency athlete performance
- PubMed: Hepcidin exercise absorption
- PubMed: Female endurance ferritin
- PubMed: Foot-strike hemolysis
- PubMed: IV iron carboxymaltose athletes
Connections
- Iron Benefits Hub
- Iron Overview
- Iron Deficiency Anemia
- Heme vs Non-Heme Iron
- Iron and Cognitive Performance
- Copper
- Hemoglobin and Ceruloplasmin
- Zinc
- Magnesium
- Vitamin C
- Vitamin B12
- Vitamin D3
- Iron Panel
- Complete Blood Count
- Fatigue
- Beef
- Salmon
- Organ Meats
- Hemochromatosis