Vitamin D3 for Immune Function & Respiratory Infection
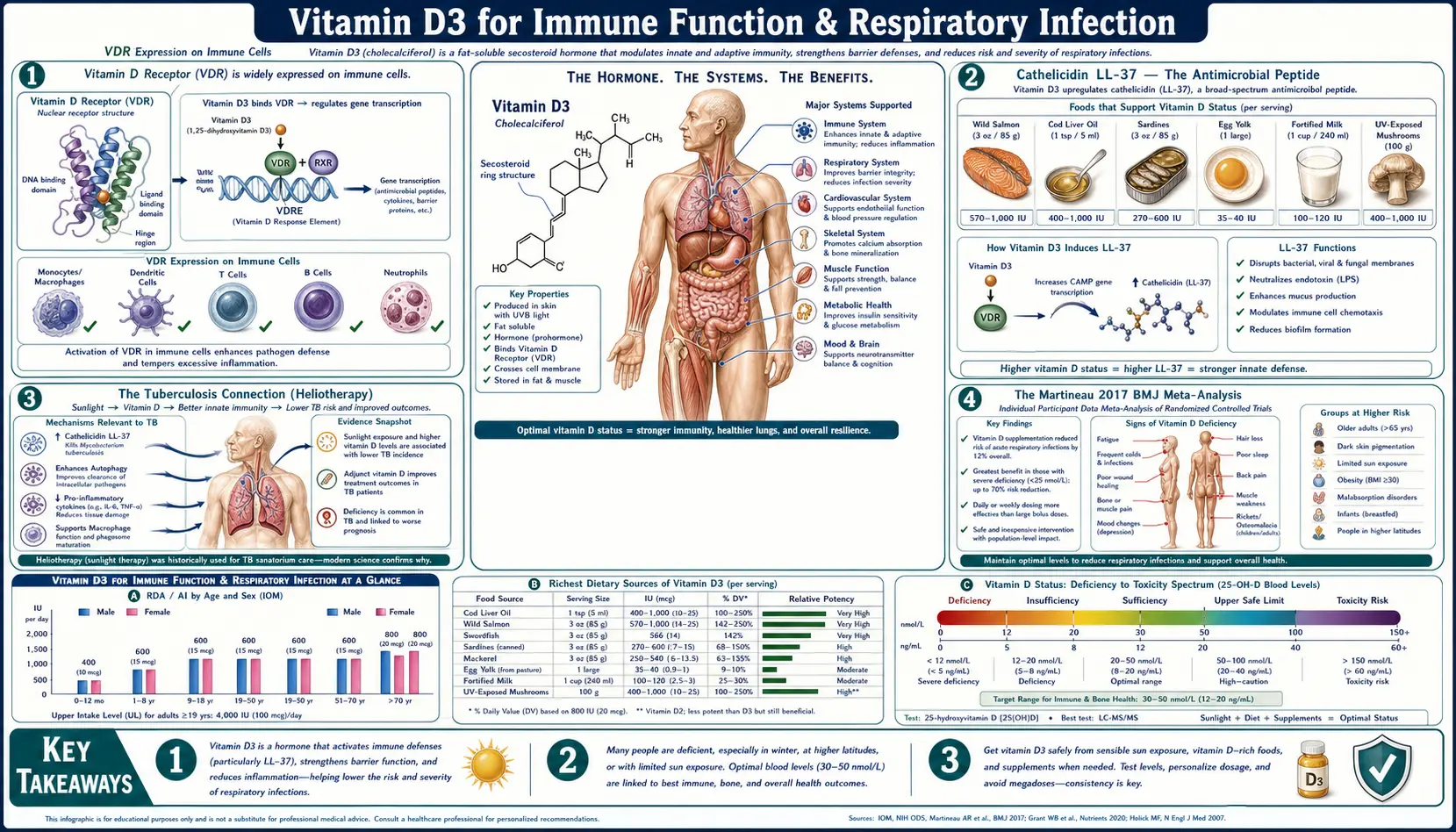
Vitamin D3 is arguably the single most important nutrient for immune defense against respiratory infection. The active form, calcitriol, is the most potent known inducer of cathelicidin LL-37 — the broad-spectrum antimicrobial peptide that macrophages secrete to kill bacteria, viruses, and fungi at the site of infection. The 2017 Martineau individual-patient-data meta-analysis in BMJ (25 RCTs, 11,321 participants) established that vitamin D supplementation reduces acute respiratory infection risk by 12% overall and by 70% in the most deficient subjects receiving daily or weekly dosing. The Castillo 2020 Cordoba pilot trial showed dramatic reduction in ICU admission and mortality when hospitalized COVID-19 patients were given calcifediol on admission. For immune outcomes, the target 25(OH)D range (50-80 ng/mL) is higher than the 30 ng/mL minimum used for bone — reflecting the higher local calcitriol production needed in immune cells to mount an effective antimicrobial response.
Table of Contents
- VDR Expression on Immune Cells
- Cathelicidin LL-37 — The Antimicrobial Peptide
- The Tuberculosis Connection (Heliotherapy)
- The Martineau 2017 BMJ Meta-Analysis
- COVID-19 Evidence: Castillo Cordoba Trial
- Th1/Th17 Modulation and Treg Expansion
- Influenza, RSV, and Other Respiratory Viruses
- Target 25(OH)D for Immune Outcomes (50-80 ng/mL)
- Bolus vs Daily Dosing — Why It Matters for Immunity
- Practical Immune-Support Protocol
- Cautions
- Key Research Papers
- Connections
- Featured Videos
VDR Expression on Immune Cells
The discovery that immune cells — not just intestine, kidney, and bone — express the vitamin D receptor (VDR) was the conceptual breakthrough that established vitamin D as far more than a bone hormone. VDR is now known to be expressed on:
- Macrophages and monocytes — the primary sites of vitamin D-induced antimicrobial peptide production
- Dendritic cells — where calcitriol modulates antigen presentation and T-cell priming
- B cells — vitamin D modulates immunoglobulin production and B-cell differentiation
- T cells (CD4+ and CD8+) — vitamin D shifts the Th1/Th2/Th17/Treg balance toward regulatory phenotypes
- NK cells — vitamin D enhances cytotoxic activity against virally infected and tumor cells
- Neutrophils — modulates chemotaxis and phagocytic capacity
Equally important: immune cells also express the activating enzyme 1α-hydroxylase (CYP27B1), meaning they can convert circulating 25(OH)D to active 1,25(OH)2D locally, independent of renal regulation. This is fundamentally different from the calcium-homeostasis circuit: in the bone axis, the kidney tightly regulates calcitriol production based on serum calcium and PTH. In the immune axis, individual macrophages crank out calcitriol on demand at the site of infection — and the supply of substrate (circulating 25(OH)D) becomes the rate-limiting step.
This is the mechanistic explanation for why immune outcomes require higher 25(OH)D levels than bone outcomes. The kidneys will generate adequate calcitriol for bone health from a modest 25(OH)D substrate pool, but local immune-cell calcitriol production is substrate-limited — macrophages at the site of an infection need plenty of 25(OH)D available to convert into the high local concentrations of calcitriol required to induce cathelicidin and other antimicrobial responses.
Cathelicidin LL-37 — The Antimicrobial Peptide
Cathelicidin (gene name CAMP, mature peptide LL-37) is a 37-amino-acid cationic antimicrobial peptide produced by macrophages, neutrophils, epithelial cells, and keratinocytes. It is a primary effector molecule of the innate immune system, with broad-spectrum activity against:
- Gram-positive bacteria (Staph aureus, Streptococcus)
- Gram-negative bacteria (E. coli, Klebsiella, Pseudomonas)
- Mycobacteria (including M. tuberculosis)
- Enveloped viruses (influenza, RSV, HIV, herpes; mechanism: membrane disruption)
- Fungi (Candida)
- Some parasites
LL-37 kills microbes by inserting into and disrupting their lipid membranes (similar to how cell-penetrating peptides work). Beyond direct killing, LL-37 also neutralizes bacterial endotoxin (lipopolysaccharide), suppressing the cytokine storm that drives sepsis pathophysiology, and modulates the recruitment of other immune cells to the infection site.
The cathelicidin (CAMP) gene contains a strong vitamin D response element (VDRE) in its promoter. When a macrophage encounters a pathogen via Toll-Like Receptor 2 (TLR2) activation, it simultaneously upregulates the VDR and 1α-hydroxylase, converts 25(OH)D to calcitriol locally, and uses that calcitriol to drive CAMP transcription — producing a burst of LL-37 specifically targeted at the site where it's needed.
This is one of the most elegant integrations of a nutrient into the immune response in human biology. It also explains the long-standing observation that vitamin D-deficient individuals are more susceptible to bacterial respiratory infections, more likely to progress to severe disease, and slower to clear infection — their macrophages simply cannot produce adequate LL-37.
The Tuberculosis Connection (Heliotherapy)
Before the discovery of antibiotics, tuberculosis was treated with two empirical interventions whose mechanism we now understand:
- Heliotherapy — sustained sun exposure at high-altitude sanatoria. Patients spent hours each day in direct sunlight, generating large amounts of cutaneous vitamin D3. Cure rates at the best European sanatoria (such as Davos in the Swiss Alps) approached 40-60% before antibiotics — far higher than untreated mortality of 50%+.
- Cod liver oil — a tablespoon a day was standard pre-antibiotic TB therapy, delivering ~1,360 IU of vitamin D3 and significant vitamin A. Studies in the early 1900s documented improved outcomes in cod-liver-oil-treated TB.
Modern molecular biology has confirmed the mechanism: M. tuberculosis-infected macrophages dramatically upregulate VDR and CYP27B1, and the resulting locally produced calcitriol drives LL-37 expression that kills intracellular mycobacteria. Vitamin D-deficient TB patients show reduced LL-37 induction and worse clinical outcomes. Several modern RCTs have shown that high-dose vitamin D adjunctive to standard anti-TB drugs accelerates sputum culture conversion (the time to a negative sputum culture, a primary efficacy outcome in TB trials) in patients with vitamin D deficiency.
The historical-medical lesson is that physicians in the pre-antibiotic era discovered empirically what molecular biology has now explained: vitamin D status profoundly affects susceptibility to and recovery from respiratory infections. The decline of sanatorium-based heliotherapy with the advent of streptomycin (1940s) meant that an entire body of clinical experience — sustained sun exposure as immune therapy — was lost from mainstream medicine for half a century.
The Martineau 2017 BMJ Meta-Analysis
Adrian Martineau and colleagues (Queen Mary, London) conducted the landmark individual-patient-data (IPD) meta-analysis that established vitamin D supplementation as a viable strategy for respiratory infection prevention. The trial-by-trial summary of 25 RCTs with 11,321 participants was published in BMJ in 2017.
Headline findings
- Overall: Vitamin D supplementation reduced acute respiratory tract infection (ARI) risk by approximately 12% (adjusted odds ratio 0.88, 95% CI 0.81-0.96)
- Daily or weekly dosing showed clear protection
- Bolus dosing (single large doses given monthly or less often) showed no benefit — an important practical finding
- The most deficient subgroup (baseline 25(OH)D < 25 nmol/L, approximately < 10 ng/mL) showed approximately 70% reduction in ARI on daily or weekly vitamin D (adjusted OR 0.30, 95% CI 0.17-0.53)
- Vitamin D-sufficient participants (baseline above 30 ng/mL) showed smaller benefit, around 25% reduction
Why this meta-analysis matters
Several features distinguish Martineau 2017 from prior unconvincing analyses:
- Individual-patient-data design — the authors obtained raw patient data from each contributing trial rather than relying on published summary statistics. This allowed subgroup analyses (baseline 25(OH)D, dosing pattern, age) that pooled aggregate data cannot perform.
- Pre-specified subgroup analyses — the dosing-pattern subgroup analysis (daily/weekly vs bolus) was pre-specified, lending credibility to the finding that bolus dosing doesn't work.
- Heterogeneity addressed — the meta-analysis revealed that earlier inconsistent results were largely explained by mixing high-bolus-dose trials (no benefit) with daily-dose trials (clear benefit). Once dosing pattern was accounted for, the picture became consistent.
The clinical takeaway from Martineau 2017 is clear: daily or weekly vitamin D supplementation in vitamin D-insufficient populations reduces respiratory infection risk meaningfully, with the greatest effect in the most deficient. Bolus dosing — despite its convenience — does not work for this indication.
COVID-19 Evidence: Castillo Cordoba Trial
The Castillo et al. 2020 pilot RCT (Journal of Steroid Biochemistry and Molecular Biology) was conducted at Reina Sofía University Hospital in Cordoba, Spain during the first wave of COVID-19. 76 hospitalized COVID-19 patients were randomized 2:1 to receive calcifediol (25-hydroxyvitamin D3) in addition to best available therapy versus best available therapy alone.
The calcifediol regimen: 0.532 mg on day 1, then 0.266 mg on days 3 and 7, then weekly.
Primary outcome: ICU admission
- Calcifediol group: 1 of 50 (2%) required ICU admission
- Control group: 13 of 26 (50%) required ICU admission
- Adjusted odds ratio: 0.03 (95% CI 0.003-0.25), p < 0.001
Mortality
- Calcifediol: 0 deaths
- Control: 2 deaths
- Sample size too small for definitive mortality conclusions, but the trend favored calcifediol
The Castillo trial drew criticism for its small size and quasi-randomization (patients were assigned by admitting day rather than fully randomized), but the effect size was so large that the result has driven multiple subsequent replication attempts. The COVIDIOL trial (also from Cordoba, n ≈ 930) and the Nogues 2021 trial (also Spanish, n ≈ 838) replicated the ICU-reduction effect in larger samples. Other trials in different settings produced mixed results, partially attributable to differences in baseline 25(OH)D, calcifediol vs cholecalciferol form, timing of administration, and severity of enrolled patients.
The mechanistic rationale is clear: COVID-19 is fundamentally a disease in which the innate immune response either contains the virus rapidly (and the patient recovers in days) or fails to contain it and the resulting dysregulated inflammatory cascade (the "cytokine storm") damages the lungs and other organs. Vitamin D's dual role of strengthening the early innate response (cathelicidin induction) and dampening the late hyperinflammatory response (Th17 suppression, Treg expansion) is precisely the pharmacology needed to shift COVID outcomes.
The Cordoba protocol used calcifediol rather than cholecalciferol because calcifediol raises 25(OH)D within hours rather than the days-to-weeks required for cholecalciferol — a critical consideration when treating an acutely ill patient.
Th1/Th17 Modulation and Treg Expansion
Vitamin D's adaptive immune effects are best characterized as regulatory rather than purely suppressive. Calcitriol acts on naive T-cells and dendritic cells to:
- Suppress excessive Th1 (pro-inflammatory) differentiation
- Suppress Th17 differentiation (the autoimmune-driving T-cell subset)
- Promote regulatory T-cell (Treg) expansion via FoxP3 induction
- Shift dendritic cells toward a tolerogenic phenotype
- Reduce production of pro-inflammatory cytokines (IL-2, IFNγ, TNFα, IL-17)
- Promote anti-inflammatory cytokine production (IL-10)
The Treg expansion is the mechanism most relevant to autoimmune disease prevention (see the Autoimmune page), but it also matters for infection: Treg expansion helps prevent the hyperinflammatory tissue damage that characterizes severe respiratory infection and sepsis. A well-regulated immune response clears infection quickly without collateral damage; a dysregulated response causes the lung damage that kills COVID-19, influenza, and bacterial pneumonia patients.
This dual role — strengthening the early innate response while dampening the late hyperinflammatory response — is unusual among immunomodulators. Most immune-supportive interventions (echinacea, elderberry, zinc) are pro-inflammatory in their action. Vitamin D is more sophisticated: it gives the immune system better tools (LL-37) early in the infection while preventing immune over-reaction late.
Influenza, RSV, and Other Respiratory Viruses
The Urashima 2010 trial (American Journal of Clinical Nutrition) randomized 334 Japanese schoolchildren to vitamin D3 1,200 IU/day or placebo from December through March. The vitamin D group had a 42% reduction in influenza A (RR 0.58, 95% CI 0.34-0.99) confirmed by viral antigen testing. There was no significant effect on influenza B in this trial.
The seasonality of influenza — peaking in mid-winter when UVB-driven D3 synthesis is at its annual nadir — was first noted by R. Edgar Hope-Simpson in the 1980s and has been substantially confirmed: northern-hemisphere influenza activity correlates inversely with population 25(OH)D levels. The "winter vitamin D nadir" hypothesis explains a significant portion of influenza seasonality that pure cold-and-crowding hypotheses cannot.
Other respiratory viruses follow similar patterns. RSV (respiratory syncytial virus) shows seasonal winter peaks and disproportionate severity in vitamin D-deficient infants. Rhinovirus (the common cold) is less seasonally dramatic but is also reduced in vitamin D-supplemented populations.
The mechanistic threads converge: vitamin D-replete individuals have macrophages and respiratory epithelial cells primed to produce LL-37 and other antimicrobial peptides, plus better-regulated T-cell responses that contain viral spread without excessive tissue damage. Vitamin D-deficient individuals lack both arms of this defense and are correspondingly more vulnerable.
Target 25(OH)D for Immune Outcomes (50-80 ng/mL)
The target 25(OH)D range for optimal immune outcomes is meaningfully higher than the target for bone outcomes:
| Indication | Target 25(OH)D Range | Rationale |
|---|---|---|
| Bone health | 40-60 ng/mL | PTH fully suppressed; fracture risk plateau |
| Immune function | 50-80 ng/mL | Substrate sufficiency for local immune-cell calcitriol production; Martineau most-deficient threshold |
| Autoimmune (Coimbra-style) | 100-200+ ng/mL | Suppression of pathogenic Th17 / autoreactive T-cells; requires specialist supervision with strict calcium-restricted diet |
The 50-80 ng/mL immune target is supported by the Martineau analysis (the largest effect at the lowest baseline; benefit extends through the > 30 ng/mL range), by population studies showing reduced infection rates at higher 25(OH)D, and by mechanistic considerations: immune-cell calcitriol production is substrate-limited, so higher circulating 25(OH)D produces higher local calcitriol at sites of infection.
For most adults, achieving the 50-80 ng/mL range requires 4,000-6,000 IU/day of vitamin D3 (with K2 and magnesium cofactors), tested every 3 months until stable then annually. Obese individuals typically need 50%-200% more.
Bolus vs Daily Dosing — Why It Matters for Immunity
One of the most clinically important findings of the Martineau 2017 meta-analysis was that bolus dosing (single large monthly or quarterly doses) does not produce respiratory infection protection, despite achieving similar long-term average 25(OH)D levels as daily dosing.
The proposed mechanism: bolus dosing creates a brief 25(OH)D spike followed by upregulation of 24-hydroxylase (CYP24A1), the catabolic enzyme that inactivates calcitriol. Over 2-4 weeks the bolus dose is broken down faster than usual, and average 25(OH)D drops back to baseline. The brief spike doesn't provide the steady substrate supply that immune cells need.
Additionally, the bolus dose may transiently disrupt the cytochrome P450 regulation of 1α-hydroxylase in immune cells, reducing local calcitriol production for several weeks after the dose despite high circulating 25(OH)D.
The practical implication: for immune outcomes, avoid 50,000 IU monthly or quarterly bolus regimens. Use daily (2,000-6,000 IU) or at most weekly dosing.
Bolus dosing remains acceptable for bone health (the bone axis is less sensitive to dosing pattern) and for rapid repletion of severe deficiency, but for ongoing immune support, daily dosing is the validated approach.
Practical Immune-Support Protocol
Maintenance (year-round)
- Vitamin D3: 4,000-6,000 IU/day (target 25(OH)D 50-70 ng/mL)
- Vitamin K2 (MK-7): 100-200 mcg/day
- Magnesium glycinate: 300-400 mg elemental at bedtime
- Vitamin A: ~5,000 IU/day from cod liver oil or food sources (synergistic with D3 in immune gene regulation)
- Zinc: 15-30 mg/day with food (cofactor for VDR function and independent antiviral effects)
- Selenium: 100-200 mcg/day (supports glutathione peroxidase, important for viral defense)
- Vitamin C: 500-1,000 mg/day
Acute illness (at first symptoms)
- Vitamin D3: 10,000-20,000 IU/day for 3-5 days (or calcifediol if available, 0.266 mg single dose then 0.266 mg every 3 days)
- Vitamin C: 1,000 mg every 2-3 hours to bowel tolerance
- Zinc: 30-50 mg/day (zinc acetate or gluconate lozenges every 2-3 hours during waking)
- Quercetin: 500 mg twice daily (acts as a zinc ionophore, increasing intracellular zinc)
- Return to maintenance D3 dose after 5-7 days
High-risk individuals (elderly, immunocompromised, healthcare workers, dark-skinned at high latitude)
- Test 25(OH)D and aim for the higher end of the immune range (60-80 ng/mL)
- Consider higher daily D3 doses (6,000-10,000 IU/day) with periodic 25(OH)D and serum calcium monitoring
- Daily dosing critical — do not substitute monthly bolus
Cautions
- Hypercalcemia — the main toxicity of very-high-dose D3. Test serum calcium and 25(OH)D every 3 months at doses above 5,000 IU/day, more frequently above 10,000 IU/day. Symptoms: nausea, polyuria, polydipsia, confusion, kidney stones.
- Sarcoidosis and granulomatous disease — macrophages in granulomas produce calcitriol without normal regulation; even modest D3 supplementation can produce hypercalcemia. Consult a specialist; calcitriol monitoring (not just 25(OH)D) may be needed.
- Bolus dosing — less effective for immune outcomes (Martineau 2017). Use daily or weekly dosing.
- Form selection in acute illness — calcifediol raises 25(OH)D in hours; cholecalciferol takes days-to-weeks. For ICU-level acute illness, calcifediol (Rayaldee or international equivalents) is preferred; for outpatient illness, high-dose cholecalciferol is acceptable.
- Drug interactions — glucocorticoids, anticonvulsants, rifampin, and ART all accelerate D3 catabolism; these patients need higher doses to achieve target levels.
- Magnesium adequacy — magnesium is required for the 25-hydroxylase and 1α-hydroxylase enzymes that activate D3. Magnesium-deficient patients show poor D3 response and may experience exacerbated muscle cramping or arrhythmias when D3 is added without magnesium.
- Vitamin A balance — vitamins A and D share RXR heterodimerization partners and regulate overlapping genes. Excess D without adequate A (or vice versa) can produce imbalanced gene expression. Cod liver oil provides both in natural ratio.
Key Research Papers
- Martineau AR, Jolliffe DA, Hooper RL, et al. (2017). Vitamin D supplementation to prevent acute respiratory tract infections: systematic review and meta-analysis of individual participant data. BMJ 356:i6583. — PubMed
- Castillo ME, Costa LME, Barrios JMV, et al. (2020). Effect of calcifediol treatment and best available therapy versus best available therapy on ICU admission and mortality among patients hospitalized for COVID-19: a pilot randomized clinical study. JSBMB 203:105751. — PubMed
- Liu PT, Stenger S, Li H, et al. (2006). Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science 311:1770-1773. (The seminal cathelicidin/macrophage paper) — PubMed
- Urashima M, Segawa T, Okazaki M, et al. (2010). Randomized trial of vitamin D supplementation to prevent seasonal influenza A in schoolchildren. Am J Clin Nutr 91:1255-1260. — PubMed
- Hewison M (2011). Vitamin D and innate and adaptive immunity. Vitamins & Hormones 86:23-62. — PubMed
- Cantorna MT, Snyder L, Lin YD, Yang L (2015). Vitamin D and 1,25(OH)2D regulation of T cells. Nutrients 7:3011-3021. — PubMed
- Aranow C (2011). Vitamin D and the immune system. J Investig Med 59:881-886. — PubMed
- Nogues X, Ovejero D, Pineda-Moncusi M, et al. (2021). Calcifediol treatment and COVID-19-related outcomes. J Clin Endocrinol Metab. — PubMed
- Bergman P, Lindh AU, Björkhem-Bergman L, Lindh JD (2013). Vitamin D and respiratory tract infections: a systematic review and meta-analysis of randomized controlled trials. PLoS One. — PubMed
- Wei R, Christakos S (2015). Mechanisms underlying the regulation of innate and adaptive immunity by vitamin D. Nutrients. — PubMed
- Martineau AR, Cates CJ, Urashima M, et al. (2016). Vitamin D for the management of asthma. Cochrane Database Syst Rev. — PubMed
- Cannell JJ, Vieth R, Umhau JC, et al. (2006). Epidemic influenza and vitamin D. Epidemiol Infect. (The Hope-Simpson hypothesis revival.) — PubMed
PubMed Topic Searches
- PubMed: vitamin D respiratory infection prevention
- PubMed: cathelicidin LL-37 vitamin D macrophage
- PubMed: vitamin D COVID-19 calcifediol
- PubMed: vitamin D influenza seasonality
- PubMed: vitamin D tuberculosis treatment
Connections
- Vitamin D3 Overview
- D3 Benefits Hub
- D3 for Bone Health
- D3 for Autoimmune Disease
- D3 for Mood & Depression
- Vitamin C
- Vitamin A
- Vitamin K2
- Zinc
- Selenium
- Magnesium
- Immune Boosting
- Echinacea
- Elderberry
- Vitamin D Test
- Infectious Disease