Immune Boosting — Vitamin D and K2
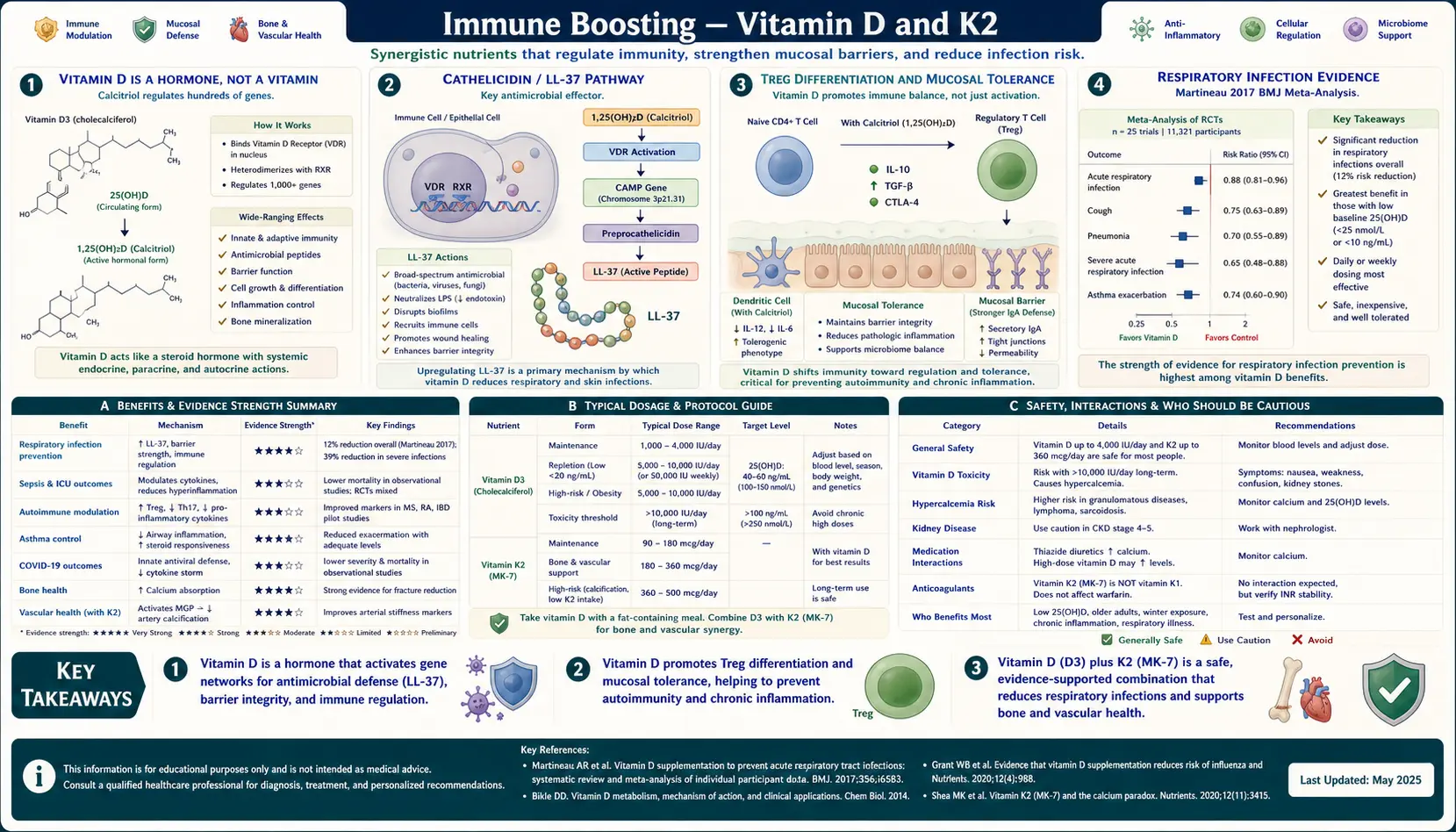
Vitamin D is the only "vitamin" the human body manufactures from sunlight, the only one with a dedicated nuclear hormone receptor (the VDR), and the only fat-soluble vitamin whose deficiency tracks latitude and winter rather than dietary intake. Its activated form — 1,25-dihydroxyvitamin D — acts as a steroid hormone, binding the VDR/RXR heterodimer to switch on hundreds of genes including the antimicrobial peptide cathelicidin (LL-37) and the regulatory transcription factor FoxP3 of Treg cells. K2 (menaquinone, MK-7 form) is the indispensable directional partner that activates matrix Gla protein, sending the calcium that D mobilizes into bone rather than into arteries, kidneys, and soft tissue. This deep-dive walks through the mechanism, the target ranges, and the practical regimen for the D / K2 axis as an immune intervention.
Table of Contents
- Vitamin D Is a Hormone, Not a Vitamin
- The Cathelicidin / LL-37 Pathway
- Treg Differentiation and Mucosal Tolerance
- The Respiratory Infection Evidence (Martineau Meta-Analysis)
- Why Latitude and Season Matter
- K2 (MK-7) as the Directional Partner
- Target Serum 25(OH)D Ranges
- Practical D + K2 Dosing Protocol
- The Magnesium Cofactor Requirement
- Cautions and Interactions
- Key Research Papers
- Connections
- Featured Videos
Vitamin D Is a Hormone, Not a Vitamin
The "vitamin" label is a historical misnomer. Vitamins are by definition substances the body cannot synthesize that must be obtained from diet. Vitamin D fails this definition on two counts: it is manufactured in skin from 7-dehydrocholesterol under ultraviolet B exposure, and its active form (1,25-dihydroxyvitamin D, calcitriol) functions identically to a steroid hormone — binding a dedicated nuclear receptor (the VDR), heterodimerizing with retinoid X receptor (RXR), and binding vitamin-D response elements (VDREs) in the promoters of hundreds of target genes.
The two-step activation is itself a regulatory feature: skin-synthesized or diet-derived cholecalciferol (D3) is hydroxylated in the liver to 25-hydroxyvitamin D (the form measured on standard blood tests), then activated in the kidney by 1-alpha-hydroxylase to the active 1,25-dihydroxyvitamin D. Critically, immune cells — macrophages, dendritic cells, activated T and B cells — also express 1-alpha-hydroxylase locally and produce 1,25-D autocrine and paracrine signaling at the site of immune activation, independent of the kidney's systemic regulation. This is why local immune effects of Vitamin D depend on having enough substrate (serum 25(OH)D) for local conversion, not just on the systemic blood level of activated hormone.
The VDR is expressed in essentially every nucleated cell type in the human body. The targets of VDR-driven transcription include genes for the antimicrobial peptides cathelicidin and beta-defensin 2, the regulatory transcription factor FoxP3 of regulatory T cells, the cytokines IL-10 and IL-37, the calcium-binding protein calbindin, and several hundred others. This breadth of action is the molecular basis for the diverse health effects associated with Vitamin D status: not just bone and calcium homeostasis, but innate antimicrobial defense, adaptive immune regulation, and modulation of risk for autoimmune disease.
The Cathelicidin / LL-37 Pathway
The cleanest molecular demonstration of Vitamin D's role in innate immunity comes from the cathelicidin pathway. Cathelicidin (gene CAMP) encodes a precursor that is proteolytically cleaved to release LL-37, a 37-residue antimicrobial peptide with broad-spectrum activity against bacteria, mycobacteria, viruses, and fungi. LL-37 punches pores in microbial membranes and also recruits immune cells via a chemokine-like signaling role.
The 2006 Liu et al. paper in Science established the modern mechanistic story. Macrophages exposed to Mycobacterium tuberculosis activate Toll-like receptor 2 (TLR2), which upregulates both the VDR and the 1-alpha-hydroxylase enzyme. The macrophage converts circulating 25(OH)D to active 1,25(OH)2D autocrinously, which then drives VDR-mediated transcription of cathelicidin. Macrophages from individuals with low serum 25(OH)D could not produce adequate LL-37 in response to TLR activation, restoring this capacity simply by adding 25(OH)D to the culture medium.
This is a direct mechanistic explanation for the longstanding empirical observation that Vitamin D deficiency correlates with increased rates of tuberculosis (used clinically before antibiotics — tuberculosis sanatoria placed patients in sunny mountain settings), respiratory tract infection, and bacterial skin infection. The pathway is also why winter, when serum 25(OH)D drops in temperate latitudes, is the season of respiratory infection — not coincidentally, but causally.
Treg Differentiation and Mucosal Tolerance
Beyond innate antimicrobial peptide induction, Vitamin D shapes adaptive immunity by promoting the development of regulatory T cells (Tregs). FoxP3 — the master transcription factor that defines and maintains the Treg lineage — has a vitamin-D response element in its promoter region. Naive CD4+ T cells encountering antigen in the presence of 1,25-dihydroxyvitamin D and TGF-beta differentiate preferentially toward the Treg phenotype rather than the Th17 phenotype.
Tregs are the brake pedal of the immune system. They produce IL-10 and TGF-beta, suppress activation of effector T cells against self-antigens and commensal microbes, and maintain mucosal tolerance to the trillions of bacteria in the gut and the food-derived antigens crossing the gut epithelium. Loss of Treg function is implicated in essentially every autoimmune disease and in many allergic and atopic conditions.
Epidemiologic data is consistent with the mechanism. Multiple sclerosis incidence rises with latitude and falls in childhood-emigration studies (children who emigrate before puberty to lower-latitude countries take on the lower MS incidence of the new country). Type 1 diabetes shows the same latitude gradient. Inflammatory bowel disease prevalence correlates inversely with sun exposure. The cytokine-tolerance mechanism through VDR-FoxP3 induction is the proposed causal explanation, and several randomized trials of Vitamin D in MS and IBD have shown modest disease-modifying effects.
For the parallel mechanism of Vitamin A on the Th17/Treg axis, see our Vitamin A for Immune Function page. Both fat-soluble vitamins signal through RXR partner heterodimers and have complementary effects on adaptive immune tolerance — one reason cod liver oil (combined A and D) has performed better in some traditional and modern protocols than isolated D alone.
The Respiratory Infection Evidence (Martineau Meta-Analysis)
The most rigorous clinical evidence for Vitamin D and immune function comes from the Martineau et al. 2017 BMJ individual-participant-data meta-analysis of 25 randomized controlled trials, pooling 10,933 participants. Key findings:
- Vitamin D supplementation reduced the risk of acute respiratory tract infection by approximately 12% across all participants
- The protective effect was substantially larger (approximately 70% reduction) in participants with baseline 25(OH)D below 25 nmol/L (10 ng/mL) — the frankly deficient subgroup
- Daily or weekly low-dose supplementation was protective; large bolus dosing (>30,000 IU at once, monthly or less frequent) was NOT protective and may have been counterproductive
- The protective effect was seen across age groups, latitude, and baseline status — though the magnitude of benefit was concentrated in deficient subjects
The take-home message from Martineau is twofold. First, Vitamin D supplementation does reduce respiratory infection risk, but the effect size matters — in adequately replete subjects, the absolute reduction is small. Second, the dosing schedule matters: physiologic daily dosing works, large pharmacologic boluses do not. This makes biologic sense — a single megadose produces transient supraphysiologic levels that feedback-suppress local immune-cell 1-alpha-hydroxylase activity, while steady daily dosing keeps serum 25(OH)D in the optimal range without inducing feedback inhibition.
Why Latitude and Season Matter
Vitamin D synthesis in skin requires UVB photons in the 290–315 nm wavelength range. The intensity of UVB at the Earth's surface depends on solar zenith angle — the higher the sun in the sky, the shorter the atmospheric path the UVB photons must traverse, and the less ozone-absorption they experience. Above approximately 40 degrees latitude (Boston, New York, Madrid, Beijing, Sydney), the solar zenith angle is too low from late October through March for meaningful Vitamin D synthesis at all. Skin can be exposed for hours and produce essentially zero Vitamin D during the winter months at these latitudes.
Even at lower latitudes, the synthesis window is limited to mid-day (roughly 10 AM to 2 PM solar time, when the sun is highest), is reduced by sunscreen, by clothing, by air pollution, by glass (window glass blocks UVB), and by skin pigmentation (melanin is an efficient UV filter; dark-skinned individuals require substantially longer exposure to make the same amount of D). The result is that even in tropical settings, modern lifestyles produce widespread Vitamin D deficiency.
The seasonal pattern of respiratory infection — the "cold and flu season" of late autumn through winter in temperate climates — tracks closely with the seasonal pattern of serum 25(OH)D. The hypothesis that this is causal, rather than coincidental with school calendars or indoor-crowding behavior, is supported by the Martineau meta-analysis showing supplementation reduces winter respiratory infections in deficient subjects. It is unlikely Vitamin D is the entire explanation for winter respiratory infections — humidity, indoor crowding, and other factors contribute — but the mechanistic and observational data strongly suggest D status is a major modifiable contributor.
K2 (MK-7) as the Directional Partner
Vitamin D's primary calcium effect is to increase calcium absorption from the gut and to mobilize calcium from bone if circulating levels drop. Vitamin K2 controls where that absorbed calcium goes. The two work as a directional pair: D loads calcium into the bloodstream; K2 sends it to bone and prevents it from depositing in arteries, kidneys, and soft tissue.
The mechanism is enzymatic. K2 is the obligate cofactor for the post-translational gamma-carboxylation of glutamic acid residues on three vitamin-K-dependent proteins relevant to calcium distribution:
- Osteocalcin — produced by osteoblasts; in its gamma-carboxylated form binds calcium and incorporates it into the hydroxyapatite matrix of bone
- Matrix Gla Protein (MGP) — produced by vascular smooth muscle cells; in its gamma-carboxylated form actively inhibits calcium deposition in arterial walls. Uncarboxylated MGP is a strong risk marker for vascular calcification
- Gla-rich protein (GRP) — broader role in soft-tissue calcium regulation
Without adequate K2, both osteocalcin and MGP remain in their inactive uncarboxylated forms. Calcium that D has mobilized then deposits where it should not — the classic outcome being vascular calcification, kidney stones, and soft-tissue mineralization, while bone density paradoxically fails to improve. This is the proposed mechanism behind the small but real signal of increased cardiovascular events seen in some Vitamin D + calcium supplementation trials without K2.
K2 exists in multiple subtypes — MK-4 (short half-life, found in animal foods, also produced from K1 by tissue conversion) and the menaquinones MK-7 through MK-13 (longer half-life, produced by bacteria, MK-7 most well-studied for supplementation). MK-7 from natto (fermented soybeans) has a serum half-life of approximately 3 days, allowing once-daily dosing to maintain steady levels. MK-4 has a half-life of only a few hours, requiring multiple daily doses to maintain levels — the reason MK-7 dominates the supplement market despite the older Japanese clinical trial literature using MK-4.
The combined Vitamin D / K2 / Vitamin A relationship is sometimes called the "triumvirate." All three fat-soluble vitamins regulate calcium and tissue mineralization through nuclear receptor and post-translational mechanisms; deficiency in any of the three subverts the function of the other two. Traditional foods carrying all three together — cod liver oil, organ meats, and fatty fish — were historically major sources, which is part of why supplementation needs to be approached as a system rather than picking isolated vitamins one at a time. See our Vitamin A Benefits hub and Vitamin K2 page for additional context.
Target Serum 25(OH)D Ranges
Reference ranges from clinical labs are misleadingly low — most labs report >20 ng/mL (50 nmol/L) as "sufficient," but this threshold is defined by bone health (preventing osteomalacia and rickets), not by the broader immune and metabolic functions of Vitamin D. The Endocrine Society's 2011 guideline, the Vitamin D Society, and most clinicians working in the integrative space target a higher range:
- < 20 ng/mL (< 50 nmol/L) — deficient; bone disease risk, severely impaired innate antimicrobial defense
- 20–30 ng/mL (50–75 nmol/L) — insufficient; adequate to prevent overt bone disease but suboptimal for immune and broader functions
- 30–50 ng/mL (75–125 nmol/L) — Endocrine Society "sufficient" range
- 50–80 ng/mL (125–200 nmol/L) — "optimal" range targeted by many integrative practitioners and many of the populations with lowest disease incidence
- > 100 ng/mL (> 250 nmol/L) — supraphysiologic; hypercalcemia risk if sustained
- > 150 ng/mL (> 375 nmol/L) — toxic; symptomatic hypercalcemia, nephrocalcinosis
The exact "optimal" target is debated and varies by source. The 50–80 ng/mL range balances the apparent benefits seen in observational cardiovascular, immune, and cognitive outcomes against the safety data showing no toxicity up to approximately 100 ng/mL. Testing serum 25(OH)D once or twice annually (end-of-summer peak and end-of-winter trough) is a reasonable cadence for anyone optimizing supplementation. The relevant assay is 25-hydroxyvitamin D, not the active 1,25-dihydroxy form — the latter is tightly homeostatically regulated and does not reflect true status.
Practical D + K2 Dosing Protocol
For an adult with serum 25(OH)D in the deficient or insufficient range, targeting the 50–80 ng/mL optimal range, a typical regimen is:
- Vitamin D3 (cholecalciferol): 2,000–5,000 IU/day with the largest meal of the day (D3 absorption is fat-dependent). Begin at 2,000 IU and retest serum 25(OH)D after 8–12 weeks; adjust up to 5,000 IU/day if level remains below target, down if level exceeds 80 ng/mL.
- K2 (MK-7 menaquinone): 100–200 mcg/day with the D3 dose. Most clinical trial dosing in cardiovascular and bone applications has used 180–360 mcg/day; 200 mcg/day is a reasonable single-product target.
- Magnesium: 200–400 mg/day as glycinate, citrate, or malate. Magnesium is the cofactor for both 25-hydroxylation in liver and 1-alpha-hydroxylation in kidney; supplementing D without magnesium reduces the conversion efficiency to active hormone.
- Vitamin A: 5,000–10,000 IU/day as retinyl palmitate if dietary intake is low (rare consumers of liver or cod liver oil typically already meet needs).
For frank deficiency (< 20 ng/mL), some clinicians use a loading phase of 50,000 IU/week for 8 weeks before transitioning to the daily maintenance dose — though the Martineau meta-analysis suggests daily dosing without a loading phase is equally effective for the immune endpoints and may avoid the feedback suppression seen with large boluses.
Combined D3 + K2 supplements are widely available and convenient. Reasonable products (no specific brand recommendation) contain D3 5,000 IU plus MK-7 100–200 mcg per softgel.
The Magnesium Cofactor Requirement
Magnesium is required as cofactor at multiple steps in Vitamin D activation and signaling. The 25-hydroxylase in liver and the 1-alpha-hydroxylase in kidney both require magnesium for catalysis. The VDR itself requires magnesium for conformational stability and DNA binding. The vitamin D binding protein (DBP) requires magnesium for proper transport function.
The practical implication: in the setting of chronic magnesium deficiency — estimated to affect roughly half of American adults given current intake patterns and the magnesium-depletion effect of common medications like proton-pump inhibitors and loop diuretics — Vitamin D supplementation produces a smaller-than-expected rise in serum 25(OH)D, and the active 1,25-dihydroxy form is produced less efficiently. Some clinicians describe patients who "won't respond" to Vitamin D supplementation; in our experience these patients are almost always magnesium-deficient and respond when magnesium is added.
Magnesium glycinate at 200–400 mg/day (elemental magnesium, not total compound weight) alongside the Vitamin D dose is the simplest protocol. Glycinate form is well-absorbed, well-tolerated, and does not produce the laxative effect of magnesium oxide or magnesium citrate. For more on the broader role of magnesium see our Magnesium page.
Cautions and Interactions
- Anticoagulant interaction (warfarin) — Vitamin K2 antagonizes warfarin's mechanism (warfarin inhibits vitamin-K-dependent clotting factor synthesis). Patients on warfarin must NOT start K2 supplementation without coordinating with their prescribing clinician for INR monitoring and dose adjustment. K2 does not interact with the newer direct oral anticoagulants (apixaban, rivaroxaban, dabigatran).
- Hypercalcemia risk in granulomatous disease — sarcoidosis, tuberculosis, lymphoma, and some other granulomatous conditions can have unregulated 1-alpha-hydroxylase activity in macrophages, producing supraphysiologic active Vitamin D from any 25(OH)D substrate. Vitamin D supplementation in these conditions requires medical supervision and may be contraindicated.
- Kidney stone history — Vitamin D increases calcium absorption; patients with calcium oxalate or calcium phosphate stones may be at increased risk with high-dose supplementation. K2 partially offsets this by directing calcium to bone, but supplementation should be approached carefully with serum calcium and 24-hour urine calcium monitoring in this population.
- Hyperparathyroidism — primary or tertiary hyperparathyroidism is a contraindication; the underlying calcium dysregulation should be addressed first.
- Thiazide diuretics — reduce urinary calcium excretion; combined with Vitamin D supplementation can produce hypercalcemia.
- Bile acid sequestrants, orlistat, mineral oil — reduce absorption of fat-soluble vitamins including D and K2. Separate dosing by at least 4 hours.
- Pregnancy — Vitamin D supplementation in pregnancy at 2,000–4,000 IU/day is safe and now generally recommended; the older 600 IU/day RDA is widely viewed as inadequate. K2 is also safe in pregnancy and lactation.
Key Research Papers
- Liu PT et al. (2006). Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science; 311(5768):1770-1773. — PubMed 16497887
- Martineau AR et al. (2017). Vitamin D supplementation to prevent acute respiratory tract infections: systematic review and meta-analysis of individual participant data. BMJ; 356:i6583. — PubMed 28202713
- Cantorna MT et al. (2015). Vitamin D, regulatory T cells, and the immune response. Nutrition Reviews; 73(7):444-453. PubMed: Cantorna 2015
- Holick MF (2007). Vitamin D deficiency. NEJM; 357(3):266-281. — Search PubMed
- Schurgers LJ et al. (2007). Vitamin K-containing dietary supplements: comparison of synthetic vitamin K1 and natto-derived menaquinone-7. Blood; 109(8):3279-3283. — PubMed 17158229
- Geleijnse JM et al. (2004). Dietary intake of menaquinone is associated with a reduced risk of coronary heart disease: the Rotterdam Study. Journal of Nutrition; 134(11):3100-3105. — PubMed 15514282
- Jeffery LE et al. (2009). 1,25-Dihydroxyvitamin D3 and IL-2 combine to inhibit T cell production of inflammatory cytokines and promote development of regulatory T cells expressing CTLA-4 and FoxP3. Journal of Immunology; 183(9):5458-5467. — PubMed 19843932
- Aranow C (2011). Vitamin D and the immune system. Journal of Investigative Medicine; 59(6):881-886. — Search PubMed
- Ginde AA et al. (2009). Association between serum 25-hydroxyvitamin D level and upper respiratory tract infection in the Third National Health and Nutrition Examination Survey. Arch Intern Med; 169(4):384-390. — PubMed 19237723
- van Ballegooijen AJ et al. (2017). The synergistic interplay between vitamins D and K for bone and cardiovascular health. Int J Endocrinol; 2017:7454376. — PubMed 29138634
- Uwitonze AM, Razzaque MS (2018). Role of magnesium in vitamin D activation and function. J Am Osteopath Assoc; 118(3):181-189. — PubMed 29480918
- Holick MF et al. (2011). Evaluation, treatment, and prevention of vitamin D deficiency: Endocrine Society clinical practice guideline. J Clin Endocrinol Metab; 96(7):1911-1930. — PubMed 21646368
PubMed Topic Searches
- PubMed: Vitamin D / cathelicidin / LL-37
- PubMed: Vitamin D / Treg / FoxP3
- PubMed: Vitamin D respiratory infection
- PubMed: K2 MK-7 / MGP / vascular calcification
- PubMed: Vitamin D / magnesium cofactor
Connections
- Immune Boosting Regimen (Main Page)
- Immune Boosting Benefits Hub
- Zinc and Selenium
- Sleep and Cortisol
- Herbal Adaptogens
- Vitamin D3
- Vitamin K2
- Vitamin A for Immune Function
- Vitamin A Benefits Hub
- Magnesium (D Cofactor)
- Calcium
- Organ Meats (Liver, Cod Liver Oil)
- Eggs (Yolk D)
- Salmon (Fatty Fish D)
- Pulmonology
- Lab Tests