Immune Boosting — Zinc and Selenium
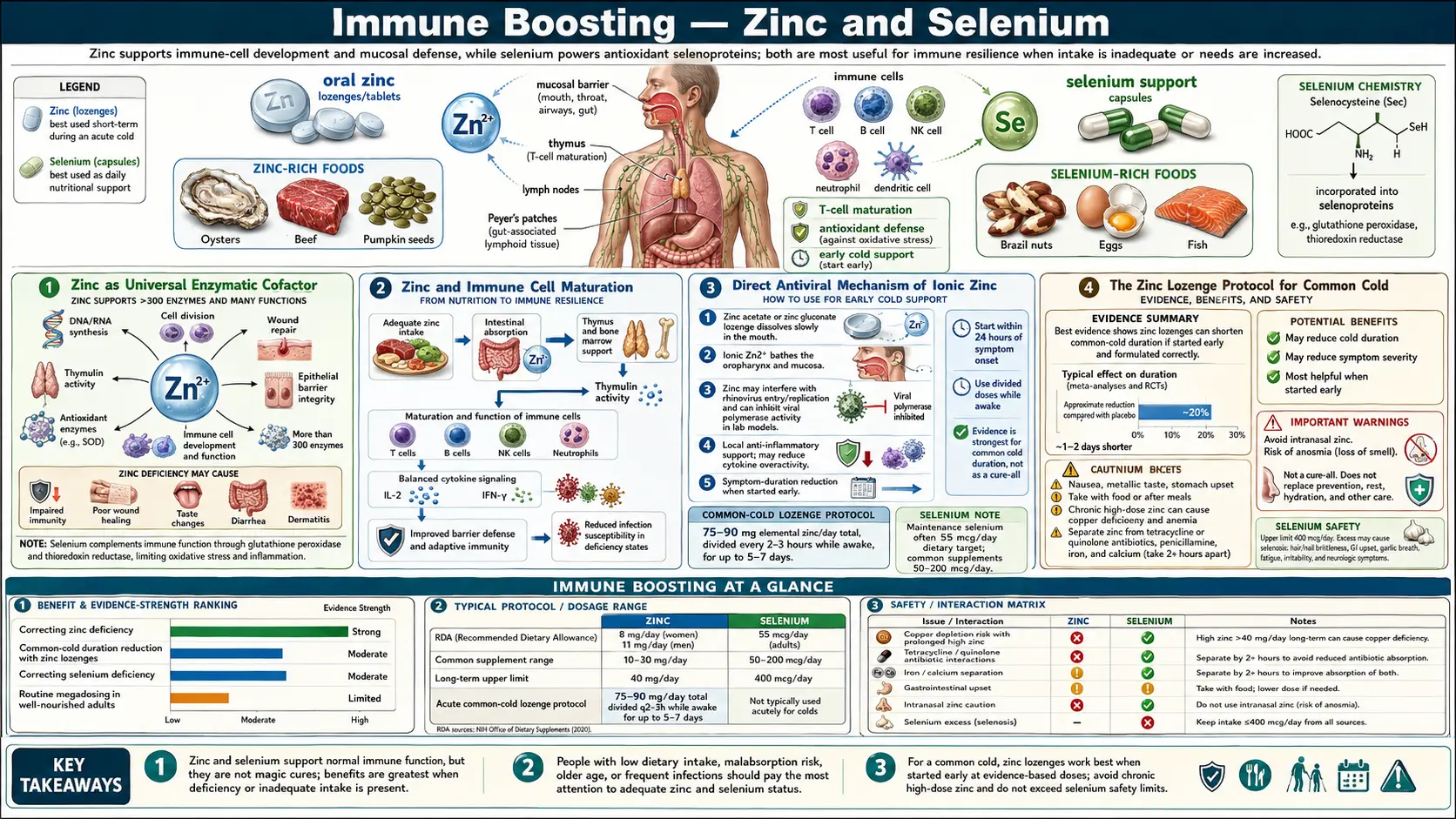
Zinc and selenium are the two redox-active trace minerals most central to antiviral and antibacterial defense. Zinc is the structural cofactor of more than 300 enzymes and several thousand zinc-finger transcription factors, the master regulator of T-cell and NK-cell maturation in the thymus, and a direct inhibitor of viral RNA-dependent RNA polymerase activity inside infected cells. Selenium is the catalytic atom in glutathione peroxidase and thioredoxin reductase — the two enzymes that determine whether oxidative stress damages or signals — and selenium status determines the in-vivo mutation rate of RNA viruses (the Beck mouse-coxsackievirus model). Together they govern both the cytotoxic capacity of innate immune cells and the antioxidant buffer that keeps that cytotoxicity from collaterally damaging host tissue. This deep-dive walks through the mechanism, the clinical evidence, and the practical dosing of both minerals as immune interventions.
Table of Contents
- Zinc as Universal Enzymatic Cofactor
- Zinc and Immune Cell Maturation
- Direct Antiviral Mechanism of Ionic Zinc
- The Zinc Lozenge Protocol for Common Cold
- Zinc and Copper Balance
- Selenium and Glutathione Peroxidase
- The Beck Mouse-Coxsackievirus Model
- Keshan Disease and Selenium Geography
- Practical Zinc and Selenium Dosing
- Cautions and Interactions
- Key Research Papers
- Connections
- Featured Videos
Zinc as Universal Enzymatic Cofactor
Zinc is the second most abundant trace mineral in the human body after iron. Total body zinc is approximately 2 to 3 grams, distributed across virtually every tissue but concentrated in skeletal muscle (60%), bone (30%), skin, liver, and the prostate. Unlike iron, zinc is not stored in a dedicated reservoir — there is no zinc equivalent of ferritin. Body zinc is functional zinc, in constant turnover, and serum zinc reflects recent intake more than total body content.
The biological role of zinc is dominated by its catalytic and structural function in enzymes. More than 300 human enzymes require a zinc atom for catalysis — including carbonic anhydrase, alkaline phosphatase, RNA polymerase, DNA polymerase, alcohol dehydrogenase, matrix metalloproteinases, and the Cu/Zn superoxide dismutase that protects against oxidative damage. Beyond enzymatic roles, zinc is the structural ion of approximately 3,000 zinc-finger transcription factors that bind DNA via tetrahedral coordination of zinc by cysteine and histidine residues. These zinc-finger transcription factors regulate roughly 10% of the human genome.
The clinical relevance: zinc deficiency simultaneously cripples hundreds of biochemical processes. The clinical phenotype of severe zinc deficiency includes growth retardation in children, hypogonadism in adolescent boys (Ananda Prasad's classic 1961 Iran cohort), dermatitis, delayed wound healing, taste and smell impairment (dysgeusia and anosmia), night blindness (zinc is required for activation of the rhodopsin visual cycle), diarrhea, and dramatically increased susceptibility to infection. The famous Prasad work in Iranian villages, where adolescents lived on unleavened bread (phytate binds zinc and prevents absorption) and had striking growth failure that reversed completely with zinc supplementation, established zinc as the prototypical "universal essential trace mineral" whose deficiency produces broad systemic phenotype.
Zinc and Immune Cell Maturation
The immune effects of zinc are concentrated at three molecular nodes: T-cell development in the thymus, natural killer cell cytotoxic function, and neutrophil chemotaxis and phagocytosis. Each depends on specific zinc-requiring proteins.
In the thymus, the hormone thymulin requires bound zinc to function. Zinc-deficient thymulin is biologically inactive. Thymulin drives the final stages of thymic T-cell maturation — the differentiation of double-positive thymocytes into mature naive CD4+ and CD8+ T cells before export to the periphery. Zinc-deficient animals develop thymic atrophy (the thymus shrinks dramatically) and have measurably reduced numbers of circulating naive T cells. Zinc supplementation in elderly subjects partially reverses age-related thymic involution and restores naive T cell output.
Natural killer cells require zinc for cytotoxic granule formation and for the conformational stability of the perforin and granzyme proteins that lyse virally infected and transformed cells. Zinc-deficient subjects have measurably reduced NK cell cytotoxic activity per cell, even when NK cell numbers are normal. The reduction in NK cytotoxicity is one mechanism by which zinc deficiency increases susceptibility to viral infection.
Neutrophils require zinc for both chemotaxis (the directed migration toward sites of infection) and phagocytosis (the engulfment of microbes). Zinc-deficient neutrophils respond more slowly to chemotactic signals and engulf fewer microbes per cell. This combines with reduced NK and T-cell function to produce the broad susceptibility-to-infection phenotype seen in zinc deficiency.
Beyond cell-autonomous effects, zinc shapes the Th1/Th2 balance through cytokine signaling. Adequate zinc supports Th1 cytokine production (IL-2, IFN-gamma), favoring cellular immunity appropriate for intracellular pathogens and viruses. Zinc deficiency shifts the balance toward Th2 (IL-4, IL-5, IL-10), favoring humoral and allergic responses inappropriately and reducing antiviral capacity.
Direct Antiviral Mechanism of Ionic Zinc
Beyond the cellular immunology effects, ionic zinc has a direct biochemical mechanism against viral replication. The 2010 te Velthuis et al. paper in PLoS Pathogens showed that Zn(2+) inhibits the RNA-dependent RNA polymerase (RdRp) of coronaviruses and arteriviruses in vitro. The mechanism appears to be allosteric — zinc binds and inhibits the catalytic activity of the polymerase, preventing replication of the viral RNA genome.
The challenge for therapeutic application is intracellular delivery. Zinc ion concentrations inside cells are tightly homeostatically regulated by zinc transporter proteins (ZnT and ZIP families); simply ingesting more zinc does not necessarily raise intracellular zinc to inhibitory concentrations. Zinc ionophores — molecules that shuttle zinc across the cell membrane independent of the regulated transporters — can raise intracellular zinc enough to achieve the antiviral effect. Quercetin (a flavonoid found in onions, apples, capers, red wine) and the antimalarial drugs chloroquine and hydroxychloroquine are zinc ionophores. The combined quercetin-plus-zinc protocol that has been used in some clinical settings exploits this ionophore mechanism.
For the upper respiratory tract specifically, ionic zinc released from zinc gluconate or zinc acetate lozenges achieves high local concentrations at the throat and nasopharyngeal mucosa — the site of rhinovirus and coronavirus replication. This local delivery is the basis for the zinc lozenge protocol covered in the next section.
The Zinc Lozenge Protocol for Common Cold
The clinical use of zinc lozenges for common cold dates to the 1984 Eby et al. randomized trial showing that zinc gluconate lozenges (23 mg elemental zinc, dissolved slowly in the mouth every 2–3 hours while awake) reduced common cold duration by an average of 7 days vs. 4 days for placebo — roughly halving the duration of symptoms when started within 24 hours of symptom onset.
Subsequent trials have produced mixed results, with the variation explained primarily by formulation. The active mechanism is local delivery of ionic zinc to the nasopharyngeal mucosa. Lozenges that release ionic zinc effectively work; lozenges that bind the zinc with sweeteners, flavoring agents, or organic acids (citric acid forms unreleasable zinc citrate, sorbitol and mannitol also bind zinc) do not work, regardless of zinc content listed on the label.
The Hemilä and Chalker 2017 systematic review pooled trials using formulations that released ionic zinc — primarily zinc acetate — and found a consistent reduction in common cold duration of approximately 33% (about 2 days shorter cold in the zinc arm vs. placebo) when started within 24 hours of symptom onset. The effect did not require dose-escalation; the standard 75–100 mg elemental zinc per day from acetate lozenges (typically 6–8 lozenges spaced through the day) was sufficient.
Practical protocol:
- Start within 24 hours of first symptom (sore throat, first sniffle)
- Use zinc acetate lozenges — the formulation with the most clinical evidence for ionic-zinc release. Cold-Eeze brand uses zinc gluconate; Zicam and several store-brand products use zinc acetate
- 13–18 mg elemental zinc per lozenge, dissolved slowly (not chewed) in the mouth every 2–3 hours while awake
- Continue for up to 7 days or until symptoms resolve
- Do NOT exceed 100 mg/day or use beyond 7–10 days — chronic high-dose zinc impairs copper absorption (see next section)
- Avoid lozenges with added citric acid, sorbitol, or mannitol (binds zinc)
Note: zinc nasal sprays and gels have been associated with permanent anosmia (loss of smell) in case reports and are NOT recommended. Only the lozenge route is supported by the clinical trial literature.
Zinc and Copper Balance
Chronic high-dose zinc supplementation produces secondary copper deficiency. The mechanism: high luminal zinc in the small intestine induces production of metallothionein, an intestinal binding protein that has higher affinity for copper than for zinc. Copper bound to metallothionein in enterocytes is sloughed off in normal turnover and lost in stool rather than absorbed. Sustained zinc intake above approximately 50 mg/day for more than a few weeks can produce measurable copper depletion, with clinical sequelae of microcytic anemia (copper is required for iron incorporation into hemoglobin), neutropenia, neuropathy, and the rare but well-documented "zinc-induced myeloneuropathy" that mimics Vitamin B12 deficiency.
The practical protocol for chronic zinc supplementation:
- If supplementing zinc at > 25 mg/day for > 4 weeks, add 1–2 mg of copper daily (as copper gluconate or bisglycinate, NOT cupric oxide which has poor bioavailability)
- Take copper at a different time of day from zinc (zinc inhibits copper absorption acutely)
- The zinc-to-copper ratio in long-term supplementation should not exceed approximately 15:1; the recommended physiologic ratio is closer to 10:1
- For acute short-term use (1-week zinc lozenge protocol for common cold), copper supplementation is not required — the duration is too short to produce copper depletion
For more on the broader role of these two minerals see our Zinc page and Copper page. The Morley Robbins work on copper, ceruloplasmin, and bioavailable iron metabolism — covered on our Morley Robbins page — is relevant for anyone exploring copper status more deeply.
Selenium and Glutathione Peroxidase
Selenium is biologically unique among trace minerals because it is incorporated into proteins as the 21st amino acid — selenocysteine — through a dedicated translation mechanism. The human genome encodes 25 known selenoproteins, of which the glutathione peroxidase family (eight isoforms, GPx1 through GPx8) and the thioredoxin reductase family (three isoforms) are the most quantitatively important.
Glutathione peroxidase catalyzes the reduction of hydrogen peroxide and organic peroxides to water and the corresponding alcohol, using glutathione (GSH) as the electron donor. This is the primary intracellular mechanism for detoxifying the hydrogen peroxide generated as a byproduct of normal aerobic metabolism and as a deliberate weapon by phagocytic immune cells. Without selenium, the GPx enzymes cannot function, and the redox balance of every cell is disrupted — oxidative damage accumulates, and the regulated use of hydrogen peroxide as a signaling molecule is also impaired.
Thioredoxin reductase reduces oxidized thioredoxin, which in turn reduces oxidized protein cysteine residues throughout the cell, maintaining the reducing intracellular environment that enzymes and signaling molecules require to function. This system is critical for both detoxification of reactive oxygen species and for the proper folding of disulfide-containing proteins.
The immune relevance: phagocytic cells (neutrophils, macrophages) use the "respiratory burst" to generate superoxide, hydrogen peroxide, and hypochlorous acid as antimicrobial weapons. Adequate selenium status allows these cells to deploy oxidative weapons against engulfed pathogens without simultaneously damaging themselves or surrounding host tissue. Selenium-deficient phagocytes either fail to mount an adequate respiratory burst (impaired pathogen killing) or undergo self-destructive oxidative damage (impaired return to homeostasis after the infection clears).
The Beck Mouse-Coxsackievirus Model
The single most striking finding in the selenium-immunity literature is the Beck et al. demonstration that selenium status affects not just the host but the virus itself. Working in the 1990s and 2000s at the University of North Carolina, Beck showed that mice deficient in selenium infected with normally benign coxsackievirus B3 developed severe viral myocarditis — and when the virus was harvested from the heart tissue of these deficient mice and re-injected into selenium-replete mice, those mice now also developed myocarditis from a virus that had previously been benign.
The mechanism: in the high-oxidative-stress environment of a selenium-deficient cell, the coxsackievirus RNA genome accumulated mutations at an accelerated rate. Specific point mutations in the viral genome converted the avirulent strain into a virulent one, and the new virulent strain was now infectious for all subsequent hosts regardless of their selenium status. This was the first clean experimental demonstration that host nutrient deficiency drives viral evolution.
The implications extend beyond coxsackievirus. The same mechanism — oxidative stress-driven viral mutation in the deficient host — has been proposed for influenza virus evolution, with some observational evidence that selenium-deficient populations may serve as "reservoir" hosts where novel influenza strains emerge. Beck's work fundamentally reshaped thinking about the interaction between host nutritional status and infectious disease epidemiology — the deficient population is not just at higher individual risk; they may be the breeding ground for the next pandemic strain.
Keshan Disease and Selenium Geography
Selenium content of crops is determined by selenium content of the underlying soil, and soil selenium varies dramatically by geography. Regions with selenium-poor soil include parts of China (most famously Keshan County in Heilongjiang Province), New Zealand, Finland, parts of central Africa, and parts of the central United States. Regions with selenium-rich soil include the western United States, parts of Venezuela, and any region built on selenium-bearing volcanic substrate.
The eponymous Keshan disease — a juvenile dilated cardiomyopathy that occurred in epidemic patterns in Keshan County, China — was traced in the 1970s to the combination of selenium deficiency in the local diet plus coxsackievirus exposure. The selenium deficiency made the local population susceptible to coxsackievirus myocarditis; the high prevalence of the virus in the population produced an endemic disease. National selenium supplementation programs in China — adding selenium to fertilizer and table salt — reduced Keshan disease incidence by >90%, validating both the mechanism and the public health intervention.
Finland implemented national selenium fortification of fertilizer in 1984 in response to recognition of widespread soil deficiency. National blood selenium levels rose from below 1 µmol/L (frankly deficient) to the modern adequate range within a few years, and the program has continued for forty years.
For Americans in the western half of the country, dietary selenium intake from wheat and other grains is typically adequate. Eastern US populations and especially those eating primarily imported or local-soil produce from selenium-poor regions may benefit from supplementation. A single Brazil nut contains approximately 70–90 micrograms of selenium — more than the daily RDA of 55 mcg — but Brazil nut selenium content is highly variable (depending on which region of Brazil the nuts came from), and overconsumption can produce selenium toxicity (selenosis: hair loss, brittle nails, garlic-breath odor, neuropathy). Two Brazil nuts daily, or 100–200 mcg/day of selenium from selenomethionine supplements, is a reasonable target.
Practical Zinc and Selenium Dosing
For routine immune-resilience supplementation in an adult:
- Zinc: 15–25 mg/day elemental zinc as picolinate, bisglycinate, or citrate. Take with food (zinc can cause nausea on empty stomach). The RDA is 11 mg/day for men, 8 mg/day for women; 15–25 mg/day is a modest excess that accounts for the variable absorption and the phytate-binding losses from grain- and legume-heavy diets.
- Copper: 1–2 mg/day as copper bisglycinate, taken at a different time of day than zinc. Required if zinc is being supplemented chronically at >25 mg/day; optional but reasonable at lower doses.
- Selenium: 100–200 mcg/day as selenomethionine (the form best incorporated into selenoproteins) or as two Brazil nuts. Stay well under the upper limit of 400 mcg/day.
For the acute zinc lozenge protocol for common cold, see the Zinc Lozenge Protocol section above. This is a short-term high-dose protocol distinct from the maintenance dosing.
Combined zinc/copper/selenium supplements are common in immune-support formulations. A reasonable starting product might contain 15 mg zinc, 1 mg copper, and 100 mcg selenium per capsule, taken with the largest meal.
Cautions and Interactions
- Zinc-induced copper deficiency — chronic zinc >50 mg/day without copper produces microcytic anemia, neutropenia, and myeloneuropathy that mimics B12 deficiency. Always add copper if supplementing zinc chronically above 25 mg/day.
- Zinc nasal sprays cause permanent anosmia — the FDA warned against Zicam Cold Remedy Nasal Gel in 2009 after >130 reports of permanent loss of smell. Lozenges are safe; nasal sprays/gels are not.
- Zinc-quinolone interaction — zinc and other divalent cations bind fluoroquinolone antibiotics (ciprofloxacin, levofloxacin) and reduce absorption substantially. Separate dosing by at least 2 hours before or 6 hours after.
- Zinc-tetracycline interaction — same chelation mechanism with tetracycline-class antibiotics (doxycycline, minocycline). Separate dosing.
- Selenium toxicity (selenosis) — sustained intake >400 mcg/day produces hair loss (most distinctive sign), brittle nails, garlic-breath odor, nausea, and neuropathy. Brazil nut overconsumption is the most common cause — some nuts contain >500 mcg each.
- Zinc and hospital infection prevention — zinc supplementation in critically ill patients in ICUs has shown modest reduction in nosocomial pneumonia in some trials. The dosing and selection considerations are specialized and should be managed by the inpatient team.
- Pregnancy — both zinc (11–12 mg/day) and selenium (60 mcg/day) are required in pregnancy at slightly higher levels than non-pregnant baseline; typical prenatal vitamins provide adequate amounts. High-dose supplementation beyond prenatal vitamin amounts should be discussed with the prenatal provider.
Key Research Papers
- Prasad AS (2008). Zinc in human health: effect of zinc on immune cells. Molecular Medicine; 14(5-6):353-357. — PubMed 18385818
- Hemilä H, Chalker E (2017). Zinc acetate lozenges for the common cold: an individual patient data meta-analysis. British Journal of Clinical Pharmacology; 82(5):1393-1398. — Search PubMed
- te Velthuis AJ et al. (2010). Zn(2+) inhibits coronavirus and arterivirus RNA polymerase activity in vitro and zinc ionophores block the replication of these viruses in cell culture. PLoS Pathogens; 6(11):e1001176. — PubMed 21079686
- Rayman MP (2012). Selenium and human health. Lancet; 379(9822):1256-1268. — Search PubMed
- Beck MA et al. (2001). Selenium deficiency and viral infection. Journal of Nutrition; 131(5):1463S-1467S. — Search PubMed
- Wessels I et al. (2017). Zinc as a gatekeeper of immune function. Nutrients; 9(12):1286. — PubMed 29186856
- Hoffmann PR, Berry MJ (2008). The influence of selenium on immune responses. Molecular Nutrition & Food Research; 52(11):1273-1280. — Search PubMed
- Read SA et al. (2019). The role of zinc in antiviral immunity. Advances in Nutrition; 10(4):696-710. — PubMed 31305906
- Avery JC, Hoffmann PR (2018). Selenium, selenoproteins, and immunity. Nutrients; 10(9):1203. — PubMed 30200430
- Eby GA, Davis DR, Halcomb WW (1984). Reduction in duration of common colds by zinc gluconate lozenges in a double-blind study. Antimicrobial Agents and Chemotherapy; 25(1):20-24. — PubMed 6367635
- Singh M, Das RR (2013). Zinc for the common cold. Cochrane Database of Systematic Reviews. — Search PubMed
- Beck MA, Levander OA, Handy J (2003). Selenium deficiency and viral infection. Journal of Nutrition; 133(5 Suppl 1):1463S-1467S. PubMed: Beck/Levander selenium
PubMed Topic Searches
- PubMed: Zinc / T cell / thymulin
- PubMed: Zinc lozenge / cold / rhinovirus
- PubMed: Selenium / selenoproteins / GPx
- PubMed: Zinc/copper balance and deficiency
- PubMed: Keshan disease / selenium
Connections
- Immune Boosting Regimen (Main Page)
- Immune Boosting Benefits Hub
- Vitamin D and K2
- Sleep and Cortisol
- Herbal Adaptogens
- Zinc
- Selenium
- Copper
- Morley Robbins Protocol
- Vitamin A for Immune Function
- Vitamin C
- Vitamin D3
- Oysters (Zinc Source)
- Beef (Zinc Source)
- Cardiology (Keshan/Selenium link)