Immune Boosting — Sleep and Cortisol
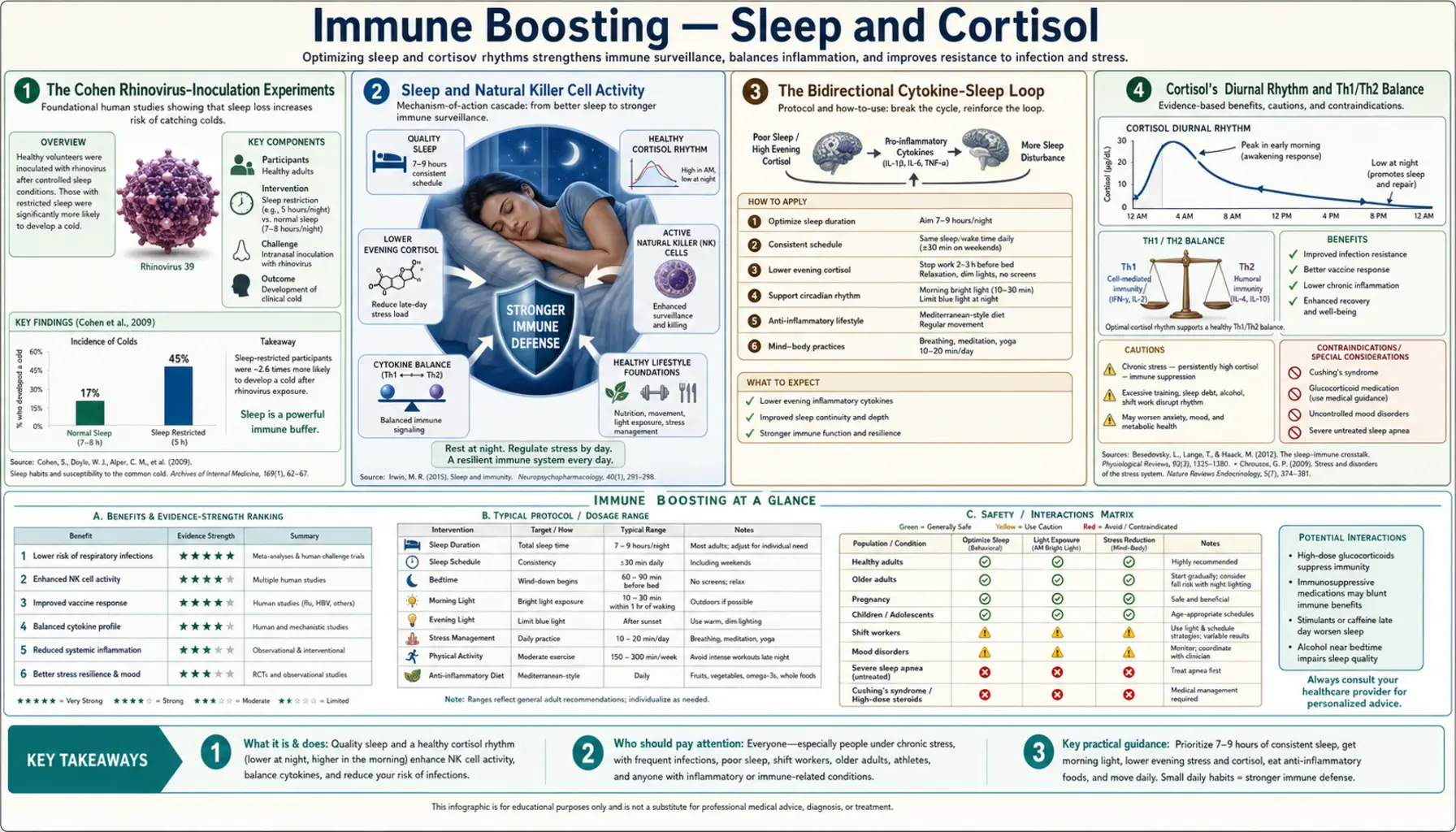
No supplement compensates for chronic sleep deprivation. A single night of restricted sleep cuts natural killer cell activity by approximately 70% (Irwin 1996), and Sheldon Cohen's rhinovirus-inoculation experiments at Carnegie Mellon showed that habitual sleepers of less than 6 hours per night had roughly four times the rate of clinical infection after standardized viral exposure compared to people sleeping 7+ hours. Cortisol, sleep, and immunity form a tight three-way loop: cortisol's diurnal rhythm sets the daily Th1/Th2 balance, sleep architecture coordinates the nightly release of immune-priming cytokines like IL-1 and TNF-alpha, and chronic HPA-axis activation suppresses lymphocyte function across the board. This deep-dive walks through the mechanism and the practical sleep-hygiene and cortisol-regulation regimen.
Table of Contents
- The Cohen Rhinovirus-Inoculation Experiments
- Sleep and Natural Killer Cell Activity
- The Bidirectional Cytokine-Sleep Loop
- Cortisol's Diurnal Rhythm and Th1/Th2 Balance
- Chronic HPA-Axis Activation
- Sleep and Vaccine Response
- Sleep Architecture: Deep Sleep and Immunity
- Practical Sleep Hygiene Protocol
- Cortisol Regulation Strategies
- Cautions and Special Populations
- Key Research Papers
- Connections
- Featured Videos
The Cohen Rhinovirus-Inoculation Experiments
The cleanest evidence that sleep affects infection risk comes from Sheldon Cohen's viral-challenge experiments at Carnegie Mellon University. The design was elegant: enroll healthy adults, measure their sleep habits over a baseline period (initially by self-report, later by actigraphy), quarantine them in a research hotel, drop a standardized inoculum of rhinovirus directly into their nasal passages, then observe over the following week which subjects developed clinical colds (defined by symptom score plus objective measures like nasal mucus weight).
The 2009 Cohen et al. paper in Archives of Internal Medicine reported the headline finding: among 153 inoculated subjects, those who had reported <7 hours of sleep per night were 2.94 times more likely to develop a clinical cold than those reporting ≥8 hours. Sleep efficiency (the fraction of time in bed actually spent asleep) was even more predictive: subjects with <92% sleep efficiency were 5.5 times more likely to develop a cold than those with ≥98% efficiency.
The 2015 Prather et al. follow-up replicated the finding with objective actigraphy measurement of sleep. Among 164 subjects inoculated with rhinovirus, those with <5 hours of objectively measured sleep had 4.5 times the rate of clinical infection compared to those with ≥7 hours, and the effect persisted after controlling for age, body composition, immune status (pre-challenge antibody titer to the inoculum virus), and stress measures.
These are some of the most rigorous cause-and-effect demonstrations in nutritional and lifestyle medicine. The viral inoculum is controlled; the outcome (clinical infection) is binary and clinically meaningful; the sleep exposure was measured before inoculation, removing reverse-causation possibility. Sleep deprivation does not just correlate with infection — it causally increases risk.
Sleep and Natural Killer Cell Activity
The mechanistic link begins with natural killer cells. NK cells are the innate-immunity sentinels that recognize and kill virally infected cells and transformed (tumor) cells. NK cytotoxic activity is measurable in vitro by harvesting NK cells from a blood draw, mixing them with a standardized tumor cell line (typically K562 cells), and measuring the percentage of target cells killed.
Irwin et al. 1996 (FASEB Journal) reported that a single night of partial sleep deprivation (3 hours of sleep instead of 8) reduced NK cell cytotoxic activity by approximately 30% the next day. A full night of total sleep deprivation reduced activity by approximately 70%. The effect was rapidly reversible — one or two nights of normal sleep restored NK activity to baseline.
The mechanism is multilayered. NK cells express receptors for both growth hormone (which surges during deep slow-wave sleep) and cortisol (which suppresses immune function and rises during sleep deprivation). The cortisol-to-growth-hormone ratio in the morning predicts NK cytotoxic activity that day. Sleep deprivation tilts the ratio toward cortisol dominance, suppressing the cells. Additionally, the diurnal trafficking of NK cells — the nightly migration from peripheral blood into lymphoid tissue and back — is disrupted by sleep loss, leaving fewer cells in the circulation where they encounter blood-borne pathogens.
The NK suppression mechanism is mechanistically connected to the Cohen rhinovirus findings. NK cells are the first line of antiviral defense in the upper respiratory mucosa. Reduced NK activity at the moment of viral exposure means the virus establishes infection more readily, and the resulting clinical infection rate is higher in sleep-restricted subjects.
The Bidirectional Cytokine-Sleep Loop
Sleep and immunity are bidirectionally coupled through cytokines. Two of the most important sleep-regulating cytokines are interleukin-1 (IL-1) and tumor necrosis factor alpha (TNF-alpha), both of which are produced by macrophages in response to pathogen recognition and both of which act on the hypothalamic preoptic area to promote slow-wave sleep.
This is why infection makes you sleepy. The same cytokines the immune system uses to signal pathogen recognition and recruit other immune cells also directly induce sleep, creating an evolutionarily adaptive feedback: when fighting infection, the body forces rest. Conversely, sleep itself induces these cytokines as a normal part of nighttime physiology — cytokine levels rise in the early hours of sleep, contributing to deep slow-wave sleep architecture, and fall during waking hours.
Sleep deprivation disrupts this regulated cytokine cycling. Chronic short sleepers have elevated baseline IL-6 and TNF-alpha during the day — a low-grade inflammatory state — combined with blunted nighttime cytokine peaks. The result is the worst of both worlds: chronic background inflammation that contributes to metabolic disease and cardiovascular risk, plus impaired acute cytokine response when actually facing infection.
The Besedovsky et al. 2019 Physiological Reviews paper is the definitive review of this sleep-immune crosstalk. It catalogues the bidirectional pathway in detail and is the standard reference for clinicians and researchers in the field.
Cortisol's Diurnal Rhythm and Th1/Th2 Balance
Cortisol is the principal glucocorticoid in humans and the dominant immunoregulatory hormone of the HPA axis. Its secretion follows a characteristic diurnal rhythm: lowest at midnight, rises sharply in the early morning hours (the cortisol awakening response peaks 30–45 minutes after waking), then declines progressively through the afternoon and evening to the midnight nadir.
This rhythm gates immune function. High morning cortisol favors Th2 (humoral) immunity and suppresses Th1 (cellular) immunity. Low evening and nighttime cortisol releases this suppression, allowing Th1 cytokine production (IL-2, IFN-gamma) to peak overnight — precisely when the body is asleep, the lymphocytes are migrating through lymphoid tissue, and the adaptive immune system is being primed for the next day's antigen encounters.
Several lines of evidence support the functional importance of this rhythm. Patients with adrenal insufficiency (Addison's disease, on hydrocortisone replacement) show altered Th1/Th2 balance and have characteristic infection susceptibility patterns. Patients on long-term high-dose glucocorticoid therapy (prednisone for autoimmune disease, asthma, or transplant) have profoundly suppressed cellular immunity and characteristic susceptibility to opportunistic infections (pneumocystis, mycobacteria, fungi). Shift workers, whose cortisol rhythm is desynchronized from environmental light cues, have measurably increased rates of infection, autoimmune disease, and cancer.
The Elenkov and Chrousos 1999 paper articulated this Th1/Th2 cortisol-shift model as a general explanation for stress-induced disease susceptibility. The model has held up well in subsequent decades of work.
Chronic HPA-Axis Activation
Acute cortisol elevation (the morning awakening response, the response to acute stress) is adaptive and immune-modulating, not immunosuppressive. Chronic HPA-axis activation — sustained elevation of cortisol from chronic psychosocial stress, chronic pain, chronic insomnia, or chronic systemic illness — is a different physiologic state and is immunosuppressive.
Glaser and Kiecolt-Glaser's 2005 Nature Reviews Immunology review synthesized 30 years of research on chronic-stress effects on immunity. The summary findings:
- Caregivers of dementia patients have measurably reduced T-cell and NK function, reduced response to vaccines, and slower wound healing
- Depression is associated with reduced T-cell mitogen responses, reduced NK activity, and elevated baseline IL-6 and TNF-alpha
- Chronic interpersonal stress (the Whitehall studies of British civil servants) predicts increased respiratory infection rates and slower recovery
- Bereavement is associated with measurably suppressed lymphocyte function for weeks following the loss
- Chronic insomnia produces a chronic-stress immunologic phenotype indistinguishable from psychosocial-stress phenotypes
The clinical implication: addressing chronic psychosocial stress, chronic insomnia, and chronic pain is part of immune optimization. Supplement regimens that ignore the chronic-stress axis are missing the dominant variable.
Sleep and Vaccine Response
If sleep deprivation suppresses immune function in general, it should also reduce the antibody response to vaccination. This has been tested directly. Spiegel et al. 2002 (JAMA) restricted sleep to 4 hours per night for 6 nights in healthy young men, then administered influenza vaccine. The sleep-restricted group developed antibody titers less than half those of the well-rested controls 10 days post-vaccination.
Lange et al. 2003 and subsequent work has shown the same pattern for hepatitis A vaccine, hepatitis B vaccine, and (more recently) SARS-related vaccines: sleep restriction in the days surrounding vaccination produces measurably blunted antibody response. The effect is large enough to be clinically meaningful, particularly for vaccines like influenza where antibody titer correlates closely with protection.
The practical implication: time vaccinations to a stretch of adequate sleep, both before and after. The recommended window is at least 7 nights of normal sleep before and 14 nights after the vaccination, to allow optimal antibody development. Travelers vaccinating before a trip should plan accordingly — vaccinating the day before a red-eye flight is biologically suboptimal.
Sleep Architecture: Deep Sleep and Immunity
Sleep is not a homogeneous state. It is composed of cyclical stages alternating between non-REM sleep (further divided into N1, N2, and N3 stages, where N3 is "slow-wave sleep" or "deep sleep") and REM sleep. The architecture of a normal night cycles through approximately 4–6 of these cycles, with slow-wave sleep predominating in the first half of the night and REM predominating in the second half.
The immune-relevant phase is slow-wave sleep. Growth hormone secretion is sharply pulsatile and concentrated in the first slow-wave sleep cycle of the night. Prolactin, which has direct immune-modulating effects, also peaks during deep sleep. The migration of naive T cells from peripheral blood into lymphoid tissue, where they encounter antigen-presenting dendritic cells, is concentrated in the deep-sleep window.
Loss of slow-wave sleep specifically — not just shortened total sleep time — reduces these immune-priming processes. The Dimitrov et al. 2019 paper in JEM showed that the rise in T-cell adhesion molecule activation during sleep depends specifically on the deep-sleep phase, and that interventions disrupting deep sleep without reducing total sleep time still impair this immune function.
Several common factors selectively reduce slow-wave sleep without necessarily reducing total time in bed: alcohol consumption (slow-wave sleep is suppressed for the first half of the night after evening alcohol), aging (slow-wave sleep declines from ~25% of sleep in adolescence to ~10% in the 60s), benzodiazepines and most non-benzodiazepine hypnotics (which produce sedation without normal sleep architecture), sleep apnea (arousals interrupt deep sleep), and chronic pain. Each of these reduces the immune-priming function of sleep even when total sleep time looks adequate on a sleep diary.
Practical Sleep Hygiene Protocol
The single most evidence-based set of interventions for sleep optimization in adults:
- Consistent sleep and wake times, including weekends — circadian regulation depends on consistency of the wake time more than the sleep time. Aim for <30 minutes variation in wake time across the week.
- Morning bright-light exposure — 10–30 minutes of outdoor light (or 10,000-lux light therapy box) within the first hour of waking. This synchronizes the suprachiasmatic nucleus master clock and produces the proper evening melatonin surge 14–16 hours later.
- Evening dim-light protocol — dim household lighting in the 2–3 hours before bed; minimize screens or use warm-color/dim modes; consider amber-tinted glasses if screens are unavoidable. Bright blue-spectrum light in the evening suppresses melatonin and delays sleep onset.
- Cool bedroom (60–67°F / 16–19°C) — sleep onset and slow-wave sleep depth depend on a falling core body temperature; cool ambient temperature facilitates this.
- No caffeine after early afternoon — caffeine has a 6-hour half-life; caffeine consumed at 3 PM is still pharmacologically active at 9 PM. Cut off intake at noon to 2 PM at the latest.
- Limit evening alcohol — alcohol shortens sleep onset but suppresses REM and slow-wave sleep for the first half of the night, producing the characteristic 3 AM awakening as it metabolizes.
- Last meal 3+ hours before bed — ongoing digestion raises core temperature and produces postprandial glucose excursions that fragment sleep.
- Bed for sleep and intimacy only — not for reading, work, TV. The bed-bedroom association with sleep is a learned cue and is degraded by other activities.
- Wind-down routine — 30–60 minutes of dim-light, low-stimulation activity (reading, gentle stretching, hot bath) signals the brain to begin the sleep-onset process.
- Address sleep apnea — if snoring, witnessed apneas, morning headache, or persistent daytime sleepiness, formal sleep study is warranted. Untreated sleep apnea fragments deep sleep and is independently immunosuppressive.
Cortisol Regulation Strategies
For HPA-axis tone, the interventions overlap with sleep optimization but include additional layers:
- Morning sunlight exposure — entrains the cortisol awakening response to occur at the right time of day and at appropriate magnitude
- Regular moderate exercise (not late evening) — exercise transiently raises cortisol but improves the overall diurnal rhythm and lowers chronic baseline. Evening high-intensity exercise can disrupt sleep onset; morning or afternoon timing is preferred
- Meditation, yoga, breathwork — activate parasympathetic nervous system tone, reducing chronic sympathetic activation. The Mindfulness-Based Stress Reduction (MBSR) protocol has substantial clinical evidence for cortisol-rhythm normalization
- Social connection — psychosocial isolation is itself a chronic stressor; regular connection with family, friends, community modulates HPA tone
- Adaptogenic herbs — particularly Ashwagandha (clinical trial evidence for cortisol reduction in chronically stressed adults; see our Herbal Adaptogens deep-dive)
- Phosphatidylserine — modest evidence for blunting cortisol response to acute stress at 100–300 mg/day; soy-derived PS is the most widely studied form
- L-theanine — 100–200 mg promotes alpha-wave brain activity and modest reduction in subjective stress; well-tolerated
Avoid: chronic high-dose caffeine (potentiates the cortisol response to stress), nightly alcohol (suppresses parasympathetic tone in the second half of the night), and screen-based work in the last hour before bed (combines blue light melatonin suppression with cognitive activation).
Cautions and Special Populations
- Shift workers — circadian misalignment is a known carcinogen (IARC Group 2A) and produces sustained immune suppression. Mitigation strategies (consistent rotation patterns, strategic light exposure, melatonin timing) help but do not fully restore normal physiology
- New parents, caregivers — chronic sleep fragmentation produces the same immune suppression as voluntary sleep restriction. Strategic napping, partnered sleep-shift sharing, and explicit recovery time are warranted
- Untreated depression and anxiety — both are associated with disrupted sleep architecture and elevated cortisol; addressing the underlying mood condition is part of the immune intervention
- Chronic pain — fragments sleep and elevates HPA tone; pain management is part of sleep optimization
- Menopause — estrogen withdrawal causes sleep fragmentation and disrupts deep-sleep architecture; addressing menopausal symptoms (including consideration of hormone replacement) is part of the immune intervention in this population
- Cushing's syndrome and exogenous glucocorticoid therapy — chronically elevated cortisol from primary disease or prescribed steroids produces severe immune suppression. Sleep hygiene alone will not normalize immune function in these settings; underlying condition must be addressed
- Sleep medications — benzodiazepines, Z-drugs (zolpidem, eszopiclone), and most over-the-counter "PM" formulations (diphenhydramine) produce sedation without normal sleep architecture. They restore sleep quantity but not necessarily sleep quality in the immune-priming sense
Key Research Papers
- Cohen S et al. (2009). Sleep habits and susceptibility to the common cold. Archives of Internal Medicine; 169(1):62-67. — PubMed 19139325
- Prather AA et al. (2015). Behaviorally assessed sleep and susceptibility to the common cold. Sleep; 38(9):1353-1359. — PubMed 26118561
- Irwin M et al. (1996). Partial night sleep deprivation reduces natural killer and cellular immune responses in humans. FASEB Journal; 10(5):643-653. — PubMed 8621064
- Besedovsky L et al. (2019). The sleep-immune crosstalk in health and disease. Physiological Reviews; 99(3):1325-1380. — PubMed 30920354
- Dhabhar FS (2014). Effects of stress on immune function: the good, the bad, and the beautiful. Immunologic Research; 58(2-3):193-210. — PubMed 24798553
- Glaser R, Kiecolt-Glaser JK (2005). Stress-induced immune dysfunction: implications for health. Nature Reviews Immunology; 5(3):243-251. — PubMed 15738954
- Lange T et al. (2010). Effects of sleep and circadian rhythm on the human immune system. Annals of the New York Academy of Sciences; 1193:48-59. — PubMed 20398008
- Spiegel K et al. (2002). Effect of sleep deprivation on response to immunization. JAMA; 288(12):1471-1472. — Search PubMed
- Dimitrov S et al. (2019). Gαs-coupled receptor signaling and sleep regulate integrin activation of human antigen-specific T cells. Journal of Experimental Medicine; 216(3):517-526. — PubMed 30755455
- Elenkov IJ, Chrousos GP (1999). Stress hormones, Th1/Th2 patterns, pro/anti-inflammatory cytokines and susceptibility to disease. Trends in Endocrinology and Metabolism; 10(9):359-368. — PubMed 10511695
- Cohen S, Tyrrell DA, Smith AP (1991). Psychological stress and susceptibility to the common cold. NEJM; 325(9):606-612. — PubMed 1713648
- Irwin MR (2015). Why sleep is important for health: a psychoneuroimmunology perspective. Annual Review of Psychology; 66:143-172. — PubMed 25061767
PubMed Topic Searches
- PubMed: Sleep deprivation / NK cells
- PubMed: Cortisol rhythm / Th1/Th2
- PubMed: Chronic stress / immunity
- PubMed: Sleep / vaccine antibody response
- PubMed: Slow-wave sleep / growth hormone / immunity
Connections
- Immune Boosting Regimen (Main Page)
- Immune Boosting Benefits Hub
- Vitamin D and K2
- Zinc and Selenium
- Herbal Adaptogens
- Ashwagandha (Cortisol Reduction)
- Rhodiola
- L-Theanine
- Vitamin D3
- Magnesium (Sleep Support)
- Endocrinology
- Neurology
- Lab Tests (Cortisol)
- All Remedies
- All Vitamins