Phosphorus for Cell Membranes
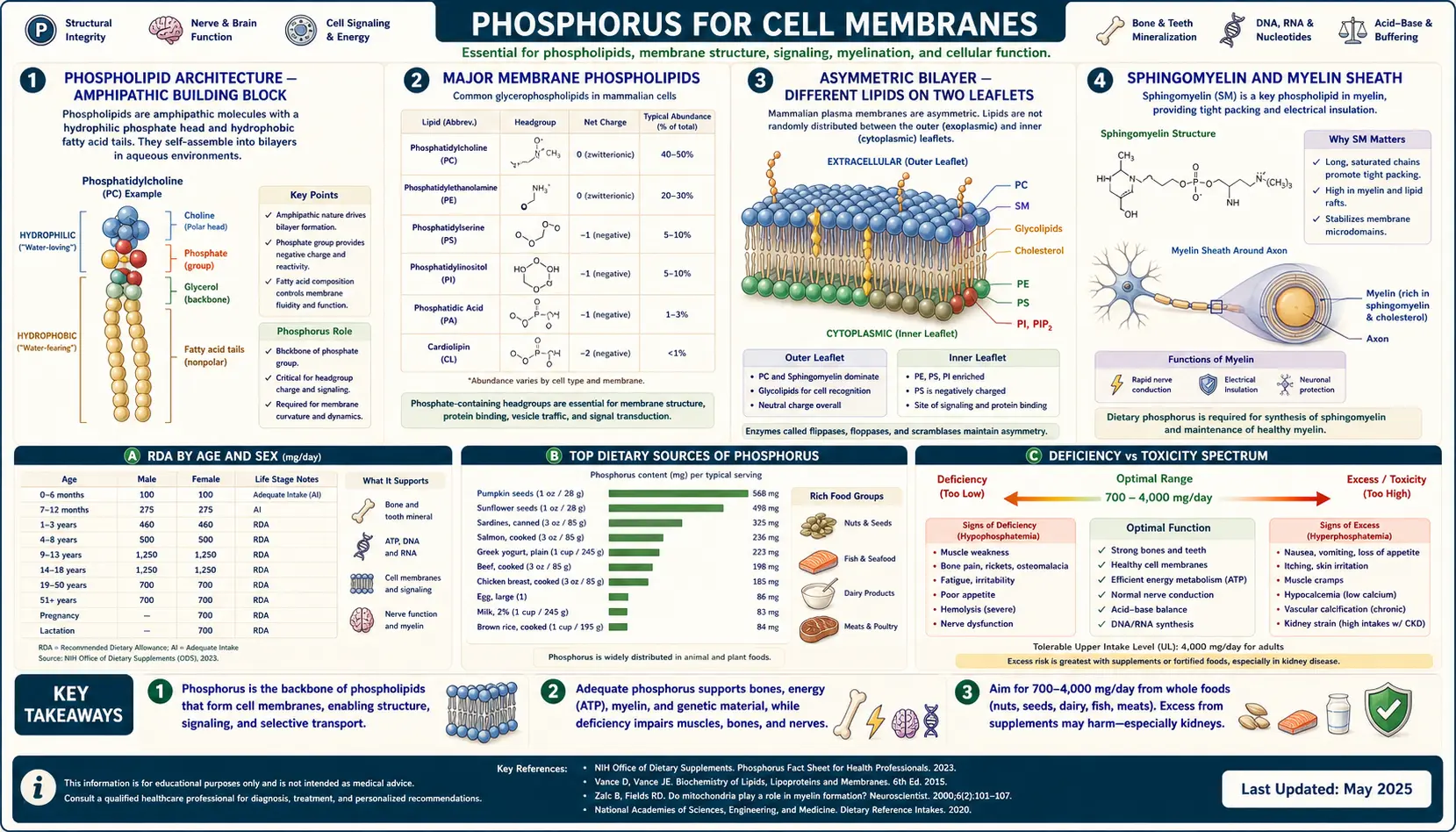
Every membrane in every cell of the human body is built from phospholipids — amphipathic molecules with a phosphate-containing polar head group and two fatty acid hydrophobic tails. The phosphate head is what makes the bilayer possible: it is the chemical handle that orients the molecule, exposes the polar surface to the aqueous interior of the cell or the extracellular fluid, and buries the hydrophobic tails away from water. Beyond their structural role, phospholipids are signaling molecules of remarkable subtlety — phosphatidylinositol-4,5-bisphosphate (PIP2) is cleaved by phospholipase C into inositol trisphosphate (IP3) and diacylglycerol (DAG), two of the most consequential second messengers in cell biology. The membrane is not just a wall; it is the cell's control surface, and phosphorus is the chemistry that makes the surface possible. This deep-dive walks through each major phospholipid class, the signaling roles, lipid raft microdomains, and the evidence on supplemental phosphatidylserine for cognition and athletic recovery.
Table of Contents
- Phospholipid Architecture — the Amphipathic Building Block
- The Major Membrane Phospholipids
- The Asymmetric Bilayer — Different Lipids on the Two Leaflets
- Sphingomyelin and the Myelin Sheath
- Phosphoinositide Signaling — PIP2, IP3, DAG, and PIP3
- Lipid Rafts and Membrane Microdomains
- Phosphatidylserine Exposure as an "Eat-Me" Signal
- Supplemental Phosphatidylserine — Evidence Review
- Phosphatidylcholine, Choline, and Cognitive Health
- Dietary Sources and Practical Recommendations
- Key Research Papers
- Connections
- Featured Videos
Phospholipid Architecture — the Amphipathic Building Block
A phospholipid is a small molecule with three modular parts: a glycerol (or sphingosine) backbone, two long fatty acid chains attached at the 1 and 2 positions, and a phosphate group attached at the 3 position with a polar head group attached to the phosphate. The two fatty acid tails are hydrophobic (water-avoiding), the phosphate plus head group is hydrophilic (water-loving). The molecule is amphipathic — it has both characteristics — and in water it spontaneously organizes into structures that bury the tails and expose the heads.
The simplest organization is a micelle (a single layer with tails inward, heads outward). For most biological purposes, however, phospholipids organize into a bilayer — two leaflets with the hydrophobic tails facing each other in the middle and the polar heads facing the aqueous environments on both sides. The bilayer is approximately 5 nm thick, fluid (lipids diffuse laterally rapidly), and forms the universal architecture of cell membranes — the plasma membrane that surrounds the cell, the inner and outer mitochondrial membranes, the endoplasmic reticulum, the Golgi apparatus, the lysosomal membrane, the nuclear envelope, and the membranes of secretory vesicles.
The phosphate group is what makes the architecture possible. Without the highly polar phosphate-plus-head-group, the molecule would not have the necessary amphipathic character. Without the negative charge of the phosphate, ion binding (especially calcium binding by phosphatidylserine) and protein recruitment to the membrane surface would not work. The chemistry of cellular life depends on phosphorus at this very basic structural level.
The Major Membrane Phospholipids
The mammalian cell membrane contains a small handful of distinct phospholipid species, each with a different head group attached to the phosphate, each with different physical properties, and each with different signaling and structural roles.
- Phosphatidylcholine (PC) — the most abundant membrane phospholipid in mammalian cells, typically 40 to 50 percent of total membrane phospholipid. The head group is choline (a quaternary amine). PC is the workhorse structural phospholipid and the primary substrate for hepatic VLDL assembly. The cell synthesizes PC from choline through the Kennedy pathway, and the precursor choline is an essential nutrient.
- Phosphatidylethanolamine (PE) — the second most abundant, typically 20 to 30 percent. The head group is ethanolamine. PE is concentrated in the inner leaflet of the plasma membrane and is particularly enriched in nervous tissue and mitochondrial membranes. It plays a role in membrane fusion and curvature because of its small head group relative to its tail volume.
- Phosphatidylserine (PS) — less abundant overall (5 to 10 percent of total membrane lipid) but disproportionately important. The head group is serine, providing both a carboxyl group and an amino group, giving PS a net negative charge at physiological pH. PS is almost entirely confined to the inner leaflet of the plasma membrane in healthy cells; its appearance on the outer leaflet is a signal of apoptosis.
- Phosphatidylinositol (PI) and its phosphorylated derivatives (PIP, PIP2, PIP3) — a small percentage of total membrane lipid (5 to 10 percent including all the phosphorylated derivatives), but the most important signaling lipid class. The head group is inositol (a six-membered sugar alcohol with five free hydroxyls available for phosphorylation in many combinations).
- Sphingomyelin (SM) — the only major phospholipid built on a sphingosine backbone rather than glycerol. The head group is phosphocholine (the same as in PC). SM is highly enriched in the myelin sheath that insulates nerve axons; it accounts for 10 to 20 percent of plasma membrane lipid in most cells and considerably more in neural tissue.
- Phosphatidylglycerol and cardiolipin — specialized phospholipids of the inner mitochondrial membrane, with cardiolipin essentially exclusive to that location. Cardiolipin is a doubly-phospholipid molecule with four fatty acid tails and is required for normal function of the electron transport chain.
Each of these has phosphorus at its structural core. None of them can be synthesized without an adequate supply of cellular phosphate, and none of them can perform their structural or signaling function without the phosphate head group.
The Asymmetric Bilayer — Different Lipids on the Two Leaflets
The plasma membrane is not symmetric. The outer leaflet (facing the extracellular space) is enriched in phosphatidylcholine and sphingomyelin. The inner leaflet (facing the cytoplasm) is enriched in phosphatidylethanolamine, phosphatidylserine, and phosphatidylinositol. The asymmetry is actively maintained by a family of ATP-dependent enzymes:
- Flippases (P4-ATPases) move PS and PE from the outer leaflet to the inner leaflet, against the concentration gradient, using ATP hydrolysis. Without active flippase function, the inner-leaflet enrichment of PS and PE would slowly equilibrate to a symmetric distribution.
- Floppases (ABC transporters) move lipids in the opposite direction, from inner to outer leaflet, also ATP-dependent.
- Scramblases (TMEM16F, others) allow rapid bidirectional movement of any lipid across the bilayer; they are activated transiently in response to calcium signals (during apoptosis, platelet activation) to break the asymmetry and expose PS on the outer leaflet.
The asymmetry matters for two reasons. First, it allows the cell to use the appearance of PS on the outer leaflet as a regulated signal (see the apoptosis section below). Second, several signaling proteins bind selectively to the negatively charged inner-leaflet lipids (PS, PIP2) through positively charged amino acid clusters or specialized lipid-binding domains; the asymmetric distribution of negative charge between the two leaflets is what allows these signaling complexes to assemble on the cytoplasmic face of the membrane and not on the extracellular face.
Sphingomyelin and the Myelin Sheath
Sphingomyelin is the dominant phospholipid of the myelin sheath, the multi-layered insulating wrap that surrounds large-diameter nerve axons in the central and peripheral nervous systems. Myelin is built by oligodendrocytes (CNS) and Schwann cells (PNS) and consists of a tightly compacted spiral of plasma membrane wrapped repeatedly around the axon. The high lipid content of myelin (approximately 70 percent of dry weight, compared with approximately 40 percent for most other membranes) is what gives white matter its appearance and its electrical insulating properties.
Sphingomyelin is particularly important because of its long, saturated fatty acid tails (often 24-carbon nervonic or lignoceric acid) and its tendency to associate with cholesterol. The combination produces a tightly packed, low-fluidity membrane that resists ion leak — exactly the property required for the saltatory conduction of nerve impulses, in which the action potential jumps electrically from one node of Ranvier to the next rather than propagating continuously down the axon.
Several inherited sphingolipid storage diseases (Niemann-Pick disease, in which a defective acid sphingomyelinase fails to break down sphingomyelin in lysosomes; metachromatic leukodystrophy; Krabbe disease) demonstrate the consequences of disrupted sphingolipid metabolism — progressive demyelination with severe neurologic disease. Even the much commoner demyelinating disorders (multiple sclerosis) involve damage to the phospholipid bilayer of myelin and impair conduction velocity through the white matter tracts of the CNS.
Phosphoinositide Signaling — PIP2, IP3, DAG, and PIP3
Phosphatidylinositol-4,5-bisphosphate (PIP2) is one of the central organizing molecules of cellular signaling. Although it makes up less than 1 percent of total membrane lipid, it sits at the crossroads of multiple signaling pathways through which extracellular signals are converted into intracellular responses.
When a G-protein-coupled receptor that activates phospholipase C-beta is stimulated — classic examples include the alpha-1 adrenergic receptor, the angiotensin II type 1 receptor, the V1 vasopressin receptor, and several muscarinic acetylcholine receptors — the phospholipase C cleaves PIP2 at the bond between the glycerol backbone and the phosphate, producing two second messengers in a single step:
- Inositol-1,4,5-trisphosphate (IP3) — a small water-soluble molecule that diffuses through the cytoplasm and binds to IP3 receptors on the endoplasmic reticulum membrane, opening calcium channels and releasing stored Ca2+ into the cytoplasm. The released calcium then triggers downstream events including smooth muscle contraction, neurotransmitter release, and activation of calcium-calmodulin-dependent kinases.
- Diacylglycerol (DAG) — remains in the membrane (it is the lipid backbone left after the head group is cleaved) and activates several isoforms of protein kinase C (PKC), which in turn phosphorylates a large list of downstream substrates including ion channels, structural proteins, and transcription factors.
The parallel pathway involves a different phospholipase: phosphatidylinositol-3-kinase (PI3K), activated by receptor tyrosine kinases (insulin receptor, growth factor receptors) and some G-protein-coupled receptors, phosphorylates PIP2 at the 3 position of the inositol ring to produce PIP3 (phosphatidylinositol-3,4,5-trisphosphate). PIP3 recruits proteins with pleckstrin homology (PH) domains to the inner leaflet of the plasma membrane, most importantly Akt (protein kinase B), which then orchestrates downstream effects on cell survival, glucose uptake, and protein synthesis through the mTOR pathway.
The PI3K / Akt / mTOR pathway is one of the most frequently mutated signaling networks in human cancer. PTEN, the tumor suppressor that dephosphorylates PIP3 back to PIP2 and shuts the pathway off, is one of the most commonly inactivated genes across cancer types. The clinical translation includes a list of cancer drugs (alpelisib for PIK3CA-mutant breast cancer, everolimus and other mTOR inhibitors for several malignancies, and rapamycin analogs in transplant immunosuppression and rare diseases). All these therapies are, at their molecular core, manipulating the cell's response to phosphorylated phosphatidylinositol — the membrane's phosphate-decorated signaling lipid.
Lipid Rafts and Membrane Microdomains
The membrane is not a uniform fluid bilayer. It is organized into small, dynamic, sterol-and-sphingolipid-enriched microdomains called lipid rafts, with distinct lipid composition and a higher packing density than the surrounding membrane. Rafts are typically 10 to 200 nm across, transient (lifetimes of seconds), but functionally critical — they concentrate specific membrane proteins (GPI-anchored proteins, palmitoylated cytoplasmic-side proteins, several growth factor receptors) into signaling platforms where the proteins can interact efficiently.
The major raft lipids are sphingomyelin and cholesterol. Their tightly packed saturated tails (sphingomyelin) and rigid sterol body (cholesterol) produce a liquid-ordered phase that segregates from the surrounding liquid-disordered phase of the rest of the membrane (rich in phosphatidylcholine with kinked unsaturated tails). The segregation is thermodynamic — the lipids self-assemble into the energetically favorable arrangement.
Rafts matter for several disease processes: HIV uses lipid rafts as entry platforms (the gp120 envelope protein and the CD4 receptor both associate with raft microdomains), several pathogens (BK virus, prion proteins) traffic preferentially through rafts, and amyloid precursor protein processing into Alzheimer-disease-associated amyloid beta occurs preferentially in raft microdomains. The membrane microarchitecture is functionally consequential, and it depends on the chemistry of the phosphate head groups (which determine which lipids can pack together) just as much as on the cholesterol content.
Phosphatidylserine Exposure as an "Eat-Me" Signal
The asymmetric distribution of phosphatidylserine to the inner leaflet of healthy cells' plasma membrane is one of the most elegant regulated signaling systems in biology. PS is held on the inner leaflet by the ATP-dependent flippase machinery described above. When a cell commits to apoptosis (programmed cell death), one of the early downstream events is the activation of caspase-cleaved scramblase enzymes and the inactivation of flippases, both of which collapse the asymmetry and expose PS on the outer leaflet within minutes.
Macrophages and other phagocytes constitutively express PS-binding receptors (TIM family, BAI1, stabilin-2) and bridge molecules (Gas6 and protein S, both of which bridge PS to phagocyte TAM-family receptors; MFG-E8, which bridges PS to integrin alpha-v-beta-3 on phagocytes). The exposed PS on the dying cell is recognized, the cell is engulfed by the phagocyte, and the entire apoptotic cell is cleared before it can release its potentially inflammatory cytoplasmic contents. The process is so efficient that under normal conditions roughly 200 billion apoptotic cells per day are cleared from the human body without any inflammation — the phagocyte's ability to recognize the exposed PS is what allows this scale of cell-death-without-tissue-damage.
Failures of apoptotic cell clearance contribute to a list of diseases. Mutations in MFG-E8 or in macrophage TAM receptors cause systemic lupus erythematosus and other autoimmune syndromes in mice (because uncleared apoptotic cells become a chronic source of nuclear autoantigen). Defects in macrophage efferocytosis (engulfment of apoptotic cells) accelerate atherosclerosis (because dying foam cells in atherosclerotic plaques are not cleared and continue to release inflammatory contents). All these processes depend, at their molecular root, on the recognition of phosphatidylserine — the phosphate-containing "eat-me" signal — on the surface of dying cells.
Supplemental Phosphatidylserine — Evidence Review
Phosphatidylserine is one of a small number of phospholipids that is sold as an oral supplement, primarily marketed for age-related cognitive decline and for athletic recovery (in particular, blunting the cortisol response to intense exercise). The bioavailability question is real and contested — oral phospholipids are partially hydrolyzed in the gut, with some absorption as intact lyso-phospholipid and significant absorption as the freed fatty acids and head group precursors. How much intact PS actually reaches neural tissue from an oral supplement is uncertain, although enrichment of brain PS pools after oral PS feeding has been documented in animal models.
The clinical evidence base:
- Cognitive decline in older adults — several small randomized trials in the 1980s and 1990s of bovine-brain-derived PS (now off the market in most countries due to BSE concerns) showed modest improvement on memory and attention measures in adults with age-associated memory impairment. Trials with soy-derived PS (the current source) have been smaller and more equivocal but some show small benefit. The FDA has allowed a qualified health claim that "limited and preliminary scientific research suggests that PS may reduce the risk of cognitive dysfunction in the elderly," with the caveat that the evidence is inconsistent.
- Athletic recovery and cortisol blunting — a series of small trials (Monteleone et al. 1992, Starks et al. 2008, Kingsley et al. 2006) reported that oral PS at 400 to 800 mg/day blunted the cortisol response to exhaustive exercise in trained men, with modest improvement in subjective fatigue and self-reported wellbeing. The mechanism is presumed to be PS modulation of the hypothalamic-pituitary-adrenal axis through poorly understood central pathways. The effect size is modest and the trials are mostly small and unreplicated at scale.
- ADHD in children — one randomized controlled trial of PS in children with ADHD reported modest improvement on parent-reported behavior measures. Generally considered insufficient evidence for clinical use.
- Alzheimer disease — small trials in mild-to-moderate Alzheimer disease have shown small short-term improvement on memory measures but no evidence of disease modification.
Practical summary: 100–300 mg/day of soy-derived PS for cognition or 400–800 mg/day for exercise-induced cortisol blunting, taken with meals to improve fat absorption. Safety profile is benign; the main side effect is mild GI upset at higher doses. Insufficient evidence to recommend routinely but reasonable as a low-risk adjunct in age-associated cognitive complaint when other modifiable factors (sleep, exercise, B12 status, thyroid function) have been addressed.
Phosphatidylcholine, Choline, and Cognitive Health
Phosphatidylcholine and its precursor choline have a separate evidence base. Choline is now classified as an essential nutrient (the FDA established an adequate intake of 425 mg/day for women and 550 mg/day for men in 1998), and the typical Western diet often falls short, especially for vegetarians and pregnant women.
Choline serves several functions:
- Phosphatidylcholine synthesis — structural substrate for the most abundant membrane phospholipid
- Acetylcholine synthesis — the major neurotransmitter at neuromuscular junctions, in the autonomic nervous system, and in cortical and hippocampal cholinergic pathways important for memory
- Methyl donor (via betaine) — choline is oxidized to betaine, which donates a methyl group to homocysteine for remethylation to methionine, intersecting with folate and B12 metabolism
- Hepatic VLDL assembly — choline deficiency is one of the established causes of fatty liver in humans (the original observation in the 1930s in malnourished infants given high-carbohydrate formulas without choline)
- Fetal brain development — choline crosses the placenta against a concentration gradient; maternal choline status during pregnancy affects offspring hippocampal development with measurable cognitive consequences in animal models
Major dietary sources include eggs (one large egg supplies approximately 150 mg of choline, mostly as phosphatidylcholine in the yolk), beef liver (the densest source), poultry, fish, and to a lesser extent legumes and cruciferous vegetables. See our Eggs page for detail on egg nutrition. Vegans and vegetarians who avoid eggs are at particular risk of low intake and should consider deliberate choline supplementation (250–500 mg/day as choline bitartrate or choline citrate), especially during pregnancy.
Dietary Sources and Practical Recommendations
Phosphorus itself is rarely deficient on any reasonable diet. The bigger nutritional questions about phospholipid health are about the building blocks and supportive nutrients rather than about phosphate per se:
- Choline (425–550 mg/day AI) — eggs, beef liver, poultry, fish, legumes
- Omega-3 fatty acids (EPA, DHA) — the fatty acid tails of membrane phospholipids are influenced by dietary fatty acid intake; chronically high omega-3 intake meaningfully increases membrane DHA content, particularly in neural tissue. Major sources include fatty fish (salmon, sardines, mackerel) and supplemental fish oil or algae oil. See our Omega-3 Fatty Acids page.
- Phosphatidylserine (PS) — primarily synthesized by the body from PE and serine; supplemental PS from soy or sunflower lecithin is available and may have modest cognitive and recovery benefits as discussed above
- Magnesium — required cofactor for the kinase enzymes that phosphorylate PI through PIP, PIP2, and PIP3. See our Magnesium page.
- B vitamins (B6, B12, folate) — methylation pathway substrates that intersect with phospholipid metabolism through the methylation of PE to PC by phosphatidylethanolamine N-methyltransferase (PEMT) in the liver
The practical implication is that the membrane health of any tissue depends on a stack of nutrients: phosphorus (rarely limiting), choline (often limiting in plant-based diets), DHA and EPA (often limiting if no fatty fish or algae oil), magnesium, and methylation-pathway B vitamins. Address the limiting nutrients in any specific individual's pattern rather than focusing on phosphorus in isolation.
Key Research Papers
- Vance JE (2015). Phospholipid synthesis and transport in mammalian cells. Traffic. — PubMed
- Balla T (2013). Phosphoinositides: tiny lipids with giant impact on cell regulation. Physiological Reviews. — PubMed
- Berridge MJ (2009). Inositol trisphosphate and calcium signalling mechanisms. BBA Molecular Cell Research. — PubMed
- Cantley LC (2002). The phosphoinositide 3-kinase pathway. Science. — PubMed
- Lingwood D, Simons K (2010). Lipid rafts as a membrane-organizing principle. Science. — PubMed
- Leventis PA, Grinstein S (2010). The distribution and function of phosphatidylserine in cellular membranes. Annual Review of Biophysics. — PubMed
- Fadok VA et al. (1992). Exposure of phosphatidylserine on the surface of apoptotic lymphocytes triggers specific recognition and removal by macrophages. J Immunology. — PubMed
- Glade MJ, Smith K (2015). Phosphatidylserine and the human brain. Nutrition. — PubMed
- Monteleone P et al. (1992). Blunting by chronic phosphatidylserine administration of the stress-induced activation of the hypothalamo-pituitary-adrenal axis in healthy men. European Journal of Clinical Pharmacology. — PubMed
- Zeisel SH, da Costa KA (2009). Choline: an essential nutrient for public health. Nutrition Reviews. — PubMed
- van Meer G et al. (2008). Membrane lipids: where they are and how they behave. Nature Reviews Molecular Cell Biology. — PubMed
- Bevers EM, Williamson PL (2016). Getting to the outer leaflet: physiology of phosphatidylserine exposure at the plasma membrane. Physiological Reviews. — PubMed
PubMed Topic Searches
- PubMed: Phospholipid bilayer and membrane structure
- PubMed: PIP2 signaling
- PubMed: PS and apoptotic clearance
- PubMed: Lipid rafts
- PubMed: Choline and cognition
Connections
- Phosphorus Overview
- Phosphorus Benefits Hub
- Phosphorus for Energy Production
- Phosphorus for Bone Mineralization
- Phosphorus for Acid-Base Balance
- Omega-3 Fatty Acids
- Magnesium
- Calcium
- Vitamin B12
- Vitamin B6
- Eggs (Choline Source)
- Organ Meats (Beef Liver)
- Salmon
- Sardines
- Cardiovascular Disease
- Neurology
- All Minerals