Molybdenum — Benefits Deep Dive
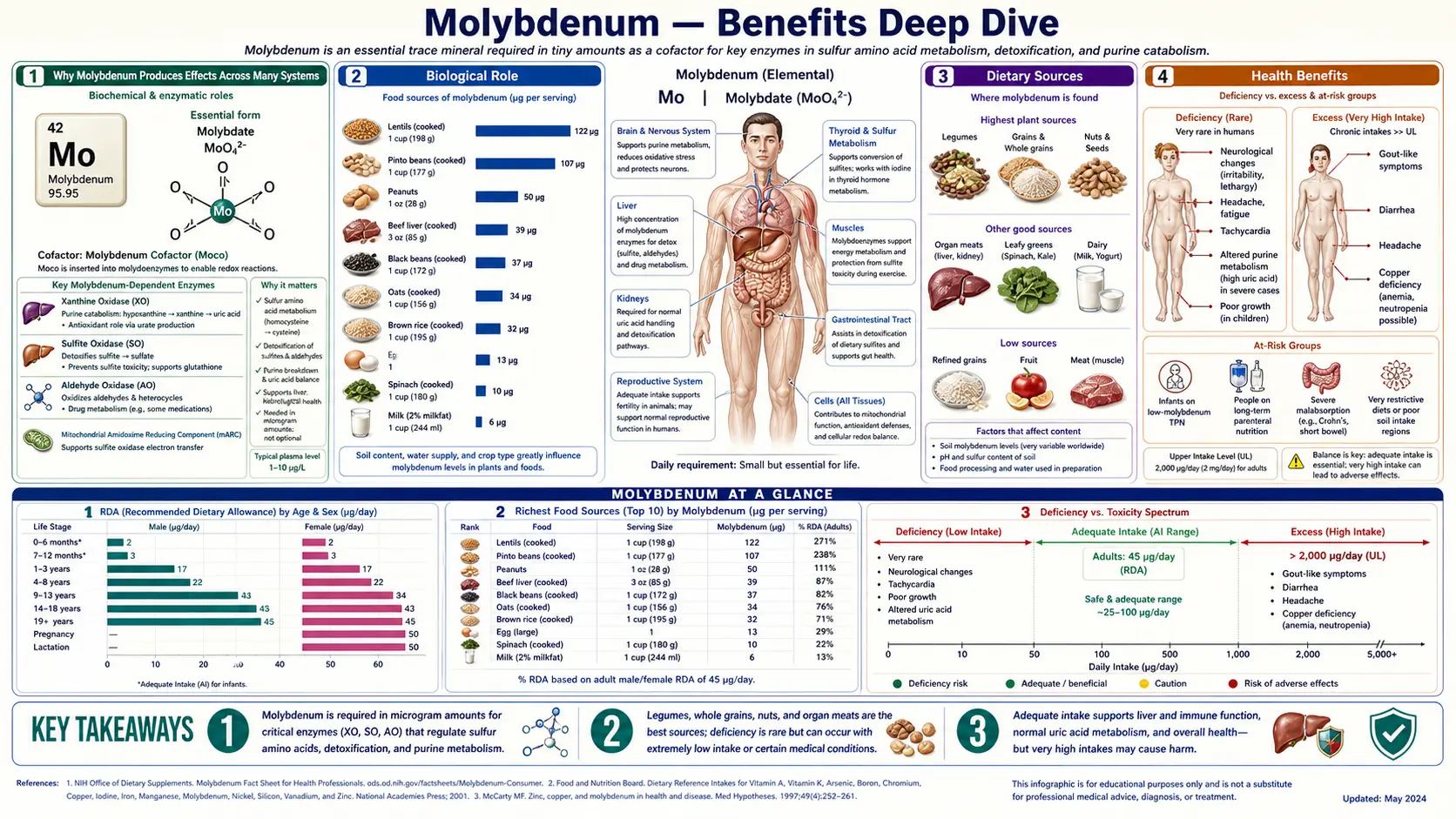
Molybdenum is the textbook example of a trace mineral that's essential despite a tiny RDA — the adult requirement is only 45 mcg per day (about one ten-thousandth of the daily protein requirement), yet without it the body cannot run four critical enzymes: sulfite oxidase (the sole enzymatic pathway for converting toxic sulfite to inert sulfate), xanthine oxidase (the terminal enzyme of purine catabolism, producing uric acid), aldehyde oxidase (a broad-spectrum oxidizer of pharmaceutical xenobiotics and environmental aldehydes), and the more recently characterized mitochondrial amidoxime reducing component (mARC). All four use the same molybdopterin cofactor — if cofactor biosynthesis fails, every molybdoenzyme fails simultaneously, producing the catastrophic neonatal disease molybdenum cofactor deficiency. The four deep-dive pages below explore the conditions where molybdenum produces the largest clinical effect — sulfite metabolism and the asthma-trigger problem, uric acid and gout pharmacology, the complex molybdenum-copper-iron triad that determines iron utilization, and the overall detoxification picture that ties the molybdoenzymes together.
Deep-Dive Articles
Detoxification
The three molybdoenzymes — sulfite oxidase, aldehyde oxidase, and xanthine oxidase — collectively provide broad-spectrum detoxification independent of the cytochrome P450 system. Covers the molybdopterin cofactor (Moco), the mitochondrial amidoxime reducing component (mARC) fourth enzyme, the catastrophic molybdenum cofactor deficiency syndrome, the breakthrough fosdenopterin (cPMP) replacement therapy for MoCD type A, and clinical relevance for adults with environmental sulfur/aldehyde exposure, mold burden, or AOX1-substrate drug metabolism considerations.
Sulfite Metabolism
Sulfite oxidase converts toxic sulfite (SO32−) to inert sulfate (SO42−) — the body's only enzymatic pathway, with no backup. Covers the molecular biology of sulfite oxidase, isolated sulfite oxidase deficiency (catastrophic pediatric phenotype), adult sulfite sensitivity (wine reactions, asthma triggers, the 1986 FDA labeling rule), the cruciferous-vegetable / high-sulfur food tolerance question, interactions with glutathione and the methylation cycle, and dietary plus supplementation strategies for sulfite-sensitive patients.
Uric Acid
Xanthine oxidase catalyzes hypoxanthine → xanthine → uric acid, the terminal two steps of purine catabolism. Covers the dual XDH/XO interconversion, uric acid as a major plasma antioxidant (50% of plasma antioxidant capacity), gout pathophysiology, allopurinol and febuxostat pharmacology (CARES trial), HLA-B*5801 risk for allopurinol hypersensitivity, the high-purine diet triggers, XO as source of reactive oxygen species in ischemia-reperfusion, hereditary xanthinuria, and tumor lysis syndrome.
Iron Utilization
Molybdenum's indirect but important role in iron metabolism via the molybdenum-copper-iron triad. High molybdenum drives copper deficiency through thiomolybdate complex formation; copper deficiency impairs the ferroxidases ceruloplasmin and hephaestin; impaired ferroxidase activity produces "iron-refractory anemia" that responds to copper, not iron. Covers aldehyde oxidase and red cell biology, the TPN case reports that revealed the connection, MOCS biosynthesis defects, and diagnostic implications when to think molybdenum.
Table of Contents
- Deep-Dive Articles
- Why Molybdenum Produces Effects Across Many Systems
- Research Papers: Detoxification & Cofactor Biology
- Research Papers: Sulfite Oxidase & Sensitivity
- Research Papers: Xanthine Oxidase & Uric Acid
- Research Papers: Aldehyde Oxidase, Copper-Iron Axis
- Research Papers: Cross-Cutting (Status, Toxicity, Drug Metabolism)
- External Authoritative Resources
- Connections
- Featured Videos
Why Molybdenum Produces Effects Across Many Systems
Most trace minerals act through a single principal mechanism — zinc activates dozens of enzymes by occupying their structural zinc sites; selenium is incorporated into selenocysteine in a small set of antioxidant enzymes; iodine is the substrate for thyroid hormone synthesis. Molybdenum is structurally unusual because it operates exclusively through one cofactor (molybdopterin, Moco) that is then used by four distinct enzymes — sulfite oxidase, xanthine oxidoreductase, aldehyde oxidase, and the more recently characterized mitochondrial amidoxime reducing component (mARC).
The shared cofactor architecture has three important consequences:
- Single point of failure — cofactor biosynthesis defects (MOCS1, MOCS2, MOCS3, GPHN mutations) simultaneously inactivate all four enzymes, producing the devastating molybdenum cofactor deficiency phenotype. The clinical picture is dominated by sulfite toxicity from sulfite oxidase failure, but the loss of xanthine and aldehyde oxidase activity compounds the damage.
- Cofactor variant sulfuration — sulfite oxidase uses the cofactor in one form (with two oxo ligands on molybdenum), while xanthine oxidase and aldehyde oxidase require a terminal sulfuration step performed by molybdenum cofactor sulfurase (MOCOS). MOCOS deficiency therefore knocks out xanthine and aldehyde oxidase activity while sparing sulfite oxidase — producing the much milder xanthinuria type II phenotype.
- Substrate breadth — from a single mineral — the same molybdenum atom (chelated by molybdopterin and held in slightly different protein scaffolds) handles sulfite, hypoxanthine, xanthine, an enormous range of aldehydes, nitrogen-containing heterocycles, N-hydroxylated drugs, and various pharmaceutical xenobiotics. The substrate range is broader than any other single trace mineral.
This is why molybdenum produces effects across systems that seem superficially unrelated — asthma triggered by preserved wine, gout attacks at the first metatarsal joint, drug pharmacokinetics for kinase inhibitors, and the copper-iron-anemia triangle. The unifying biology is the molybdopterin cofactor at the active sites of four enzymes handling biochemically distinct but mechanistically similar oxidation reactions.
The therapeutic complication is that molybdenum interacts mechanically with two other trace minerals:
- Copper — thiomolybdate complexes formed from molybdate and sulfide bind copper with extremely high affinity, depleting the copper pool. This is the basis for tetrathiomolybdate therapy in Wilson disease and the agricultural problem of molybdenum-induced copper deficiency in cattle. Practical implication: high-dose molybdenum supplementation must be balanced with adequate copper intake.
- Iron — the copper depletion just described impairs the copper-dependent ferroxidases (ceruloplasmin, hephaestin) that load iron onto transferrin. The cascade molybdenum → copper → iron explains why "iron-refractory anemia" sometimes responds to copper repletion (and indirectly to molybdenum balance) rather than to more iron. See the Iron Utilization page for the full mechanism.
Research Papers: Detoxification & Cofactor Biology
- Hille R, Hall J, Basu P (2014). The mononuclear molybdenum enzymes. Chemical Reviews. — PubMed
- Schwarz G, Mendel RR, Ribbe MW (2009). Molybdenum cofactors, enzymes and pathways. Nature. — PubMed
- Mendel RR, Bittner F (2006). Cell biology of molybdenum. Biochimica et Biophysica Acta. — PubMed
- Schwahn BC et al. (2015). Efficacy and safety of cyclic pyranopterin monophosphate in severe molybdenum cofactor deficiency type A. The Lancet. — PubMed
- Reiss J, Hahnewald R (2011). Molybdenum cofactor deficiency: mutations in GPHN, MOCS1, and MOCS2. Human Mutation. — PubMed
- Veldman A et al. (2010). Successful treatment of molybdenum cofactor deficiency type A with cPMP. Pediatrics. — PubMed
- Kisker C, Schindelin H, Rees DC (1997). Molybdenum-cofactor-containing enzymes: structure and mechanism. Annual Review of Biochemistry. — PubMed
- Schwarz G, Belaidi AA (2013). Molybdenum in human health and disease. Metal Ions in Life Sciences. — PubMed
- Rajagopalan KV, Johnson JL (1992). The pterin molybdenum cofactors. Journal of Biological Chemistry. — PubMed
- Mendel RR (2013). The molybdenum cofactor. Journal of Biological Chemistry. — PubMed
Research Papers: Sulfite Oxidase & Sensitivity
- Tan WH et al. (2005). Isolated sulfite oxidase deficiency: a case report with a novel mutation and review. Pediatrics. — PubMed
- Bender D et al. (2019). Impaired mitochondrial maturation of sulfite oxidase in a patient with severe SUOX deficiency. Human Molecular Genetics. — PubMed
- Vincent SH et al. (1991). Sulfite-induced asthma. Annals of Allergy. — PubMed
- Vally H, Misso NL (2012). Adverse reactions to the sulphite additives. Gastroenterology and Hepatology from Bed to Bench. — PubMed
- Simon RA (1998). Update on sulfite sensitivity. Allergy. — PubMed
- Belaidi AA, Schwarz G (2013). Metal insertion into the molybdenum cofactor. Biochemical Journal. — PubMed
- Touati G et al. (2000). Molybdenum cofactor deficiency: from genotype to phenotype. Journal of Inherited Metabolic Disease. — PubMed
- Stipanuk MH, Ueki I (2011). Dealing with methionine/homocysteine sulfur: cysteine metabolism to taurine and inorganic sulfur. Journal of Inherited Metabolic Disease. — PubMed
- Bouchireb K et al. (2014). Three new cases of sulfite oxidase deficiency. Pediatric Nephrology. — PubMed
- Garcia-Castello FJ et al. (2007). Cysteine sulfinic acid decarboxylase, hypotaurine and taurine. Amino Acids. — PubMed
Research Papers: Xanthine Oxidase & Uric Acid
- Harrison R (2004). Physiological roles of xanthine oxidoreductase. Drug Metabolism Reviews. — PubMed
- Choi HK et al. (2004). Purine-rich foods, dairy and protein intake, and the risk of gout in men. NEJM. — PubMed
- White WB et al. (2018). Cardiovascular Safety of Febuxostat or Allopurinol in Patients with Gout (CARES). NEJM. — PubMed
- Becker MA et al. (2005). Febuxostat compared with allopurinol in patients with hyperuricemia and gout. NEJM. — PubMed
- Hung SI et al. (2005). HLA-B*5801 allele as a genetic marker for severe cutaneous adverse reactions caused by allopurinol. PNAS. — PubMed
- Ames BN et al. (1981). Uric acid provides an antioxidant defense in humans. PNAS. — PubMed
- FitzGerald JD et al. (2020). 2020 American College of Rheumatology Guideline for the Management of Gout. Arthritis & Rheumatology. — PubMed
- Granger DN (1988). Role of xanthine oxidase and granulocytes in ischemia-reperfusion injury. American Journal of Physiology. — PubMed
- Coiffier B et al. (2008). Guidelines for the management of pediatric and adult tumor lysis syndrome. JCO. — PubMed
- Choi HK, Curhan G (2008). Soft drinks, fructose consumption, and the risk of gout in men. BMJ. — PubMed
Research Papers: Aldehyde Oxidase, Copper-Iron Axis
- Pryde DC et al. (2010). Aldehyde oxidase: an enzyme of emerging importance in drug discovery. Journal of Medicinal Chemistry. — PubMed
- Garattini E, Terao M (2012). The role of aldehyde oxidase in drug metabolism. Expert Opinion on Drug Metabolism & Toxicology. — PubMed
- Abumrad NN et al. (1981). Amino acid intolerance during prolonged TPN reversed by molybdate therapy. AJCN. — PubMed
- Brewer GJ et al. (2006). Tetrathiomolybdate for the treatment of Wilson disease. Archives of Neurology. — PubMed
- Hellman NE, Gitlin JD (2002). Ceruloplasmin metabolism and function. Annual Review of Nutrition. — PubMed
- Halfdanarson TR et al. (2008). Hematological manifestations of copper deficiency. European Journal of Haematology. — PubMed
- Anderson GJ, Frazer DM (2017). Current understanding of iron homeostasis. AJCN. — PubMed
- Ichida K et al. (2012). Identification of MOCOS gene mutations in xanthinuria type II. Human Molecular Genetics. — PubMed
- Suttle NF (2010). Copper-molybdenum-sulfur interactions in livestock nutrition. CABI. — PubMed
- Williams DM (1983). Copper deficiency in humans. Seminars in Hematology. — PubMed
Research Papers: Cross-Cutting (Status, Toxicity, Drug Metabolism)
- Turnlund JR (2002). Molybdenum metabolism and requirements in humans. Metal Ions in Biological Systems. — PubMed
- Novotny JA, Peterson CA (2018). Molybdenum. Advances in Nutrition. — PubMed
- Mendel RR, Schwarz G (2011). Molybdenum cofactor biosynthesis in plants and humans. Coordination Chemistry Reviews. — PubMed
- IOM (2001). Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc. National Academies Press. — PubMed
- Vyskocil A, Viau C (1999). Assessment of molybdenum toxicity in humans. Journal of Applied Toxicology. — PubMed
- Wuebbens MM, Rajagopalan KV (1995). Investigation of the early steps of molybdopterin biosynthesis. JBC. — PubMed
- Mendel RR, Kruse T (2012). Cell biology of molybdenum in plants and humans. Biochimica et Biophysica Acta. — PubMed
- Schwarz G (2016). Molybdenum cofactor and human disease. Current Opinion in Chemical Biology. — PubMed
- Hille R (2013). The molybdenum oxotransferases and related enzymes. Dalton Transactions. — PubMed
- Veldman A et al. (2017). Fosdenopterin restores molybdenum cofactor biosynthesis in MoCD type A. Molecular Genetics and Metabolism. — PubMed
External Authoritative Resources
- Linus Pauling Institute — Molybdenum Micronutrient Information Center — the single most authoritative scientific summary, regularly updated
- NIH Office of Dietary Supplements — Molybdenum Fact Sheet (Health Professionals)
- IOM/NASEM — Dietary Reference Intakes for Molybdenum and Trace Minerals (2001)
- UniProt — Human Sulfite Oxidase (SUOX)
- UniProt — Human Xanthine Dehydrogenase / Oxidase (XDH)
- UniProt — Human Aldehyde Oxidase 1 (AOX1)
- FDA — Nulibry (fosdenopterin) Prescribing Information
- MedlinePlus Genetics — Molybdenum Cofactor Deficiency
- PubMed — All research on molybdenum enzymes in humans
Connections
- Molybdenum (Main Page)
- Molybdenum for Detoxification
- Molybdenum for Sulfite Metabolism
- Molybdenum for Uric Acid
- Molybdenum for Iron Utilization
- All Minerals
- Copper
- Sulfur
- Selenium
- Zinc
- Methionine
- Cysteine
- N-Acetylcysteine (NAC)
- Glutathione
- Gout
- Asthma
- Uric Acid Test
- Sulfites
- Liver Cleansing
- Gerson Detoxification
- Copper-Iron Dysregulation
- Organ Meats