Alanine — Benefits Deep Dive
Alanine is the most efficient gluconeogenic amino acid in human physiology and the unique nitrogen shuttle that lets peripheral tissues (especially exercising skeletal muscle) safely export their nitrogen waste to the liver for urea-cycle disposal — the elegant biochemistry of the Cahill (glucose-alanine) cycle. Its alpha-carbon isomer beta-alanine is an entirely different molecule, the rate-limiting precursor for muscle carnosine, an intramuscular pH buffer that prolongs high-intensity exercise capacity. Alanine's namesake liver enzyme — alanine aminotransferase (ALT) — is the single most-ordered marker of hepatocyte injury on every standard liver-panel blood test in the world. Four benefit pages below explore the conditions where alanine produces the largest clinical effect — the glucose-alanine cycle and blood-sugar maintenance, beta-alanine and high-intensity exercise capacity, immune-cell amino-acid requirements during sepsis and critical illness, and ALT as the canonical liver biomarker together with the NAFLD-related elevations now affecting roughly a quarter of the global adult population.
Deep-Dive Articles
Gluconeogenesis & Blood Sugar
The glucose-alanine cycle (Cahill cycle) as the physiological link between muscle protein breakdown and hepatic glucose output, alanine aminotransferase (ALT) as the rate-limiting transaminase, liver gluconeogenesis during overnight fasting and prolonged starvation, the 60-90% increase in alanine output from exercising muscle, and alanine's role as the universal nitrogen carrier from peripheral tissues to the liver where the urea cycle disposes of the amino group safely as urea.
Beta-Alanine for Endurance
The fundamentally different beta-isomer: beta-alanine is the rate-limiting precursor for carnosine (beta-alanyl-L-histidine) in skeletal muscle, carnosine functions as the principal intramuscular pH buffer during anaerobic exertion, the harmless paraesthesia ("tingle") effect from histidine-rich receptor activation, the Harris/Hoffman 4-6 g/day loading protocol over 4-12 weeks, and the high-intensity exercise capacity gains documented across hundreds of randomized trials.
Immune Function
Alanine as a substrate for lymphocyte proliferation, immune cell amino acid requirements during activation, the glutamine-alanine relay that exchanges nitrogen-rich glutamine for nitrogen-stripped alanine in activated lymphocytes, conditional essentiality during sepsis and critical illness, and the use of intravenous alanine-glutamine dipeptides (Dipeptiven) in modern critical-care nutrition.
Liver Function
Alanine aminotransferase (ALT) as the canonical biochemical marker of hepatocyte injury on every standard liver panel, normal hepatic alanine metabolism, the alanine load test as a research tool for assessing hepatic functional reserve, non-alcoholic fatty liver disease (NAFLD) and its global ALT-elevation epidemic, drug-induced liver injury patterns, and the AST:ALT ratio in distinguishing alcoholic from non-alcoholic liver disease.
Table of Contents
- Deep-Dive Articles
- Why Alanine Produces Effects Across Many Systems
- Research Papers: Gluconeogenesis & Blood Sugar
- Research Papers: Beta-Alanine for Endurance
- Research Papers: Immune Function
- Research Papers: Liver Function
- Research Papers: Cross-Cutting (Mechanism, Status, Safety)
- External Authoritative Resources
- Connections
- Featured Videos
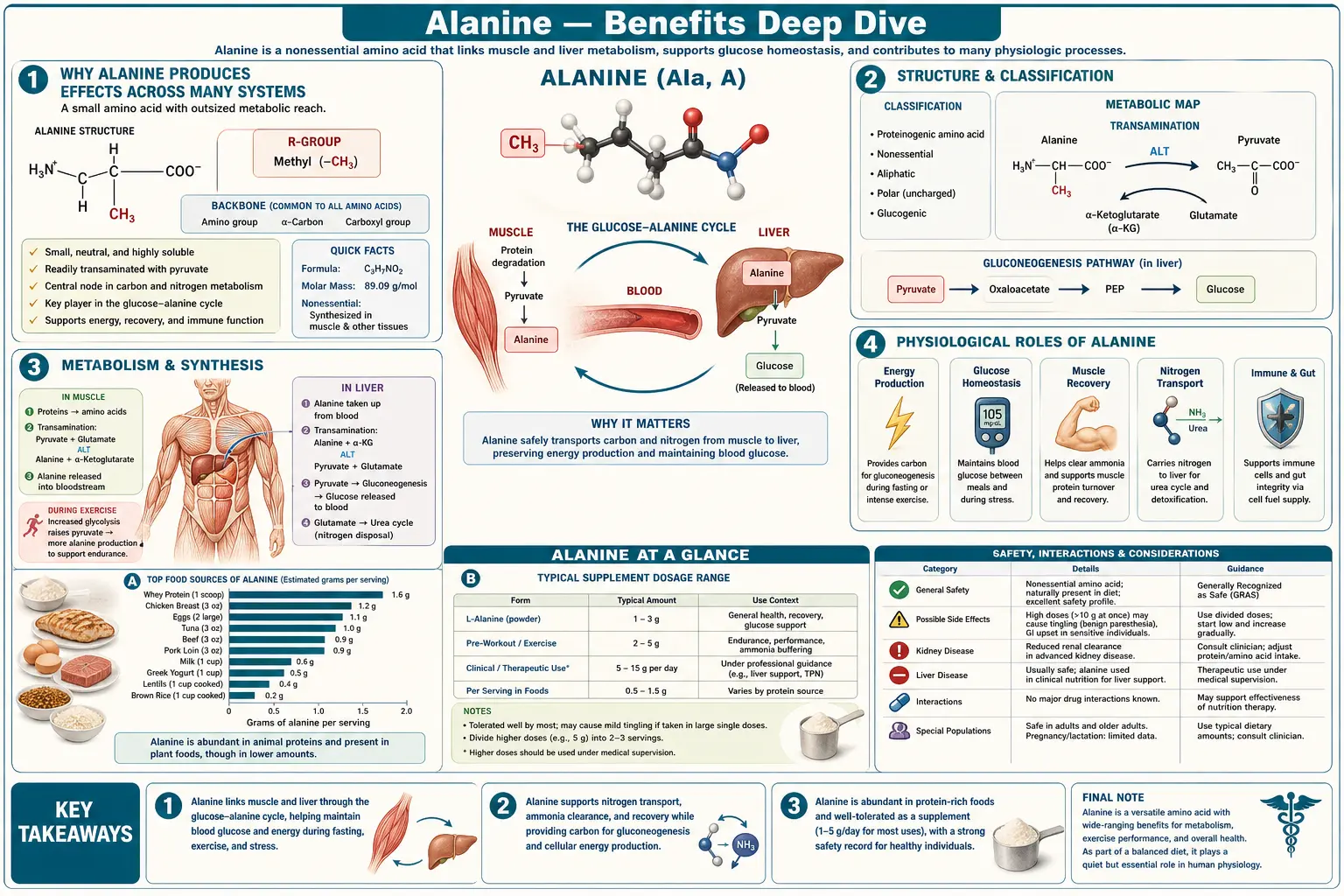
Why Alanine Produces Effects Across Many Systems
Most amino acids serve a single principal role: as a substrate in protein synthesis. Alanine is unusual because it operates simultaneously as a metabolic intermediate, a nitrogen shuttle, and the parent molecule for an entirely distinct beta-isomer with its own non-overlapping biology. Each role maps to a distinct category of clinical effect.
- The most efficient gluconeogenic amino acid — Alanine is the preferred carbon skeleton for hepatic gluconeogenesis. The reaction is structurally trivial — one transamination step converts alanine to pyruvate, which feeds directly into the gluconeogenic pathway. No other amino acid converts to glucose so cleanly or in such high flux. This is why alanine carries the bulk of inter-organ glucose precursor flow during fasting and exercise, and why glucagon secretion is amino-acid-responsive specifically to alanine.
- Universal nitrogen shuttle from periphery to liver — Skeletal muscle catabolizes branched-chain amino acids (leucine, isoleucine, valine) for local energy production during exercise and fasting, but the liberated nitrogen cannot stay in muscle (muscle lacks a complete urea cycle). The nitrogen is repackaged onto pyruvate by alanine aminotransferase (ALT) to make alanine, exported to the bloodstream, and disposed of safely by the hepatic urea cycle. This is the basis of the glucose-alanine (Cahill) cycle, and a closely parallel glutamine-alanine relay operates inside activated immune cells.
- Beta-alanine: a structurally distinct, non-proteinogenic isomer — Although named after L-alanine, beta-alanine (3-aminopropanoic acid) is not used in protein synthesis. Its sole known biological role is as the rate-limiting substrate for carnosine (beta-alanyl-L-histidine) synthesis in skeletal muscle. Carnosine functions as the principal intramuscular pH buffer during high-intensity anaerobic exercise. Oral beta-alanine supplementation can roughly double intramuscular carnosine concentration over 4-12 weeks of loading. This is the basis of beta-alanine's well-documented high-intensity exercise capacity benefits — an effect achievable through no other dietary route.
- The diagnostic enzyme that bears its name — Alanine aminotransferase (ALT, historically SGPT — serum glutamic-pyruvic transaminase) catalyzes the reversible transfer of an amino group between alanine and alpha-ketoglutarate, producing pyruvate and glutamate. ALT concentrations are extremely high inside hepatocytes and low elsewhere, which makes serum ALT the single most useful biochemical marker of liver-cell injury on every standard liver panel. ALT elevation patterns are the gateway to all liver disease diagnosis, from drug-induced hepatitis to NAFLD to viral hepatitis to autoimmune hepatitis to hereditary hemochromatosis.
The therapeutic complication is that these four roles can pull in different directions. Dietary L-alanine intake supports glucose-alanine cycling and immune cell function but does almost nothing for muscle carnosine (very little L-alanine is converted to beta-alanine in vivo). Beta-alanine supplementation builds muscle carnosine but does not contribute meaningfully to gluconeogenic substrate pools. Elevated ALT is almost always a sign of hepatocyte damage rather than an alanine surplus — it is the enzyme leaking from injured cells, not the substrate being used. And alanine-glutamine dipeptide infusions in critical care are pharmacologically distinct from oral amino-acid supplements both in kinetics and in indication. Modern best practice treats these as four genuinely different therapeutic conversations sharing only a name.
Research Papers: Gluconeogenesis & Blood Sugar
- Felig P (1973). The glucose-alanine cycle. Metabolism. — PubMed
- Felig P, Pozefsky T, Marliss E, Cahill GF (1970). Alanine: key role in gluconeogenesis. Science. — DOI: 10.1126/science.167.3920.1003
- Felig P, Marliss E, Pozefsky T, Cahill GF (1970). Amino acid metabolism in the regulation of gluconeogenesis in man. American Journal of Clinical Nutrition. — PubMed
- Cahill GF (2006). Fuel metabolism in starvation. Annual Review of Nutrition. — DOI: 10.1146/annurev.nutr.26.061505.111258
- Wahren J, Felig P, Ahlborg G, Jorfeldt L (1971). Glucose metabolism during leg exercise in man. JCI. — DOI: 10.1172/JCI106683
- Ahlborg G, Felig P, Hagenfeldt L, Hendler R, Wahren J (1974). Substrate turnover during prolonged exercise in man. JCI. — DOI: 10.1172/JCI107889
- Genuth SM, Castro J (1974). Effect of oral alanine administration in fasting obese subjects. Metabolism. — PubMed
- Owen OE, Felig P, Morgan AP, Wahren J, Cahill GF (1969). Liver and kidney metabolism during prolonged starvation. JCI. — DOI: 10.1172/JCI106013
- Snell K (1980). Muscle alanine synthesis and hepatic gluconeogenesis. Biochemical Society Transactions. — PubMed
- Petersen KF, Dufour S, Shulman GI (2005). Decreased insulin-stimulated ATP synthesis in elderly. PLOS Medicine. — DOI: 10.1371/journal.pmed.0020233
- Rothman DL, Magnusson I, Katz LD, et al. (1991). Quantitation of hepatic glycogenolysis and gluconeogenesis in fasting humans (NMR). Science. — DOI: 10.1126/science.1948033
- Garber AJ, Karl IE, Kipnis DM (1976). Alanine and glutamine synthesis and release from skeletal muscle. JBC. — PubMed
Research Papers: Beta-Alanine for Endurance
- Harris RC, Tallon MJ, Dunnett M, et al. (2006). The absorption of orally supplied beta-alanine and its effect on muscle carnosine synthesis in human vastus lateralis. Amino Acids. — DOI: 10.1007/s00726-006-0299-9
- Hoffman J, Ratamess NA, Faigenbaum AD, et al. (2008). Short-duration beta-alanine supplementation increases training volume and reduces subjective feelings of fatigue. Nutrition Research. — DOI: 10.1016/j.nutres.2007.11.004
- Trexler ET, Smith-Ryan AE, Stout JR, et al. (2015). International Society of Sports Nutrition position stand: Beta-alanine. JISSN. — DOI: 10.1186/s12970-015-0090-y
- Saunders B, Elliott-Sale K, Artioli GG, et al. (2017). Beta-alanine supplementation to improve exercise capacity and performance: a systematic review and meta-analysis. British Journal of Sports Medicine. — DOI: 10.1136/bjsports-2016-096396
- Hill CA, Harris RC, Kim HJ, et al. (2007). Influence of beta-alanine supplementation on skeletal muscle carnosine concentrations and high intensity cycling capacity. Amino Acids. — DOI: 10.1007/s00726-006-0364-4
- Stout JR, Cramer JT, Mielke M, et al. (2006). Effects of twenty-eight days of beta-alanine and creatine monohydrate supplementation on the physical working capacity at neuromuscular fatigue threshold. JSCR. — DOI: 10.1519/R-18525.1
- Boldyrev AA, Aldini G, Derave W (2013). Physiology and pathophysiology of carnosine. Physiological Reviews. — DOI: 10.1152/physrev.00039.2012
- Derave W, Ozdemir MS, Harris RC, et al. (2007). Beta-Alanine supplementation augments muscle carnosine content and attenuates fatigue during repeated isokinetic contraction bouts in trained sprinters. JAP. — DOI: 10.1152/japplphysiol.00795.2007
- Decombaz J, Beaumont M, Vuichoud J, et al. (2012). Effect of slow-release beta-alanine tablets on absorption kinetics and paresthesia. Amino Acids. — DOI: 10.1007/s00726-011-1169-7
- Sale C, Saunders B, Harris RC (2010). Effect of beta-alanine supplementation on muscle carnosine concentrations and exercise performance. Amino Acids. — DOI: 10.1007/s00726-009-0443-4
Research Papers: Immune Function
- Newsholme P, Curi R, Pithon-Curi TC, et al. (1999). Glutamine metabolism by lymphocytes, macrophages, and neutrophils: its importance in health and disease. JNB. — DOI: 10.1016/S0955-2863(99)00045-7
- Calder PC (2006). Branched-chain amino acids and immunity. Journal of Nutrition. — DOI: 10.1093/jn/136.1.288S
- Newsholme EA, Crabtree B, Ardawi MS (1985). Glutamine metabolism in lymphocytes: its biochemical, physiological, and clinical importance. QJEP. — PubMed
- Wischmeyer PE (2008). Glutamine: mode of action in critical illness. Critical Care Medicine. — DOI: 10.1097/CCM.0b013e318168ec55
- Heyland DK, Dhaliwal R, Day AG, et al. (2013). REducing Deaths due to OXidative Stress (REDOXS): glutamine and antioxidant supplementation in critically ill patients. NEJM. — DOI: 10.1056/NEJMoa1212722
- Mittendorfer B, Volpi E, Wolfe RR (2001). Whole body and skeletal muscle glutamine metabolism in healthy subjects. AJPEM. — DOI: 10.1152/ajpendo.2001.280.2.E323
- Krebs HA (1980). Glutamine metabolism in the animal body. In: Glutamine: Metabolism, Enzymology, and Regulation. — PubMed
- Curi R, Newsholme P, Procopio J, et al. (2007). Glutamine, gene expression, and cell function. Frontiers in Bioscience. — DOI: 10.2741/2197
- Wischmeyer PE, Dhaliwal R, McCall M, et al. (2014). Parenteral glutamine supplementation in critical illness: a systematic review. Critical Care. — DOI: 10.1186/cc13836
- Karinch AM, Pan M, Lin CM, Strange R, Souba WW (2001). Glutamine metabolism in sepsis and infection. Journal of Nutrition. — DOI: 10.1093/jn/131.9.2535S
Research Papers: Liver Function
- Pratt DS, Kaplan MM (2000). Evaluation of abnormal liver-enzyme results in asymptomatic patients. NEJM. — DOI: 10.1056/NEJM200004273421707
- Kwo PY, Cohen SM, Lim JK (2017). ACG Clinical Guideline: Evaluation of abnormal liver chemistries. American Journal of Gastroenterology. — DOI: 10.1038/ajg.2016.517
- Newsome PN, Cramb R, Davison SM, et al. (2018). Guidelines on the management of abnormal liver blood tests. Gut. — DOI: 10.1136/gutjnl-2017-314924
- Younossi ZM, Koenig AB, Abdelatif D, et al. (2016). Global epidemiology of nonalcoholic fatty liver disease — meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology. — DOI: 10.1002/hep.28431
- Cohen JC, Horton JD, Hobbs HH (2011). Human fatty liver disease: old questions and new insights. Science. — DOI: 10.1126/science.1204265
- Sookoian S, Pirola CJ (2012). Alanine and aspartate aminotransferase and glutamine-cycling pathway in NAFLD. World J Gastroenterology. — DOI: 10.3748/wjg.v18.i29.3775
- Cohen DE, Anania FA, Chalasani N (2006). An assessment of statin safety by hepatologists. American Journal of Cardiology. — DOI: 10.1016/j.amjcard.2005.12.010
- Prati D, Taioli E, Zanella A, et al. (2002). Updated definitions of healthy ranges for serum alanine aminotransferase levels. Annals of Internal Medicine. — DOI: 10.7326/0003-4819-137-1-200207020-00006
- Mofrad P, Contos MJ, Haque M, et al. (2003). Clinical and histologic spectrum of nonalcoholic fatty liver disease associated with normal ALT values. Hepatology. — DOI: 10.1053/jhep.2003.50229
- Williams ALB, Hoofnagle JH (1988). Ratio of serum aspartate to alanine aminotransferase in chronic hepatitis — relationship to cirrhosis. Gastroenterology. — DOI: 10.1016/0016-5085(88)90192-1
- Cohen JA, Kaplan MM (1979). The SGOT/SGPT ratio — an indicator of alcoholic liver disease. Digestive Diseases and Sciences. — DOI: 10.1007/BF01324898
Research Papers: Cross-Cutting (Mechanism, Status, Safety)
- Alanine aminotransferase (ALT) tissue distribution and clinical interpretation — PubMed
- Pyridoxal-5-phosphate (vitamin B6) as the cofactor for ALT transamination — PubMed
- D-alanine, gut microbiome, and bacterial peptidoglycan biology — PubMed
- Alanine-glutamine dipeptide (Dipeptiven) in parenteral nutrition — PubMed
- Branched-chain amino acid transamination and alanine synthesis in muscle — PubMed
- Plasma amino acid profiles and metabolic phenotyping (metabolomics) — PubMed
- Alanine and insulin secretion (pancreatic beta cells) — PubMed
- Carnosine pharmacology beyond exercise (anti-glycation, neuroprotection) — PubMed
- Branched-chain amino acid catabolism disorders (MSUD) and alanine accumulation — PubMed
- Hyperalaninemia and pyruvate carboxylase deficiency — PubMed
External Authoritative Resources
- Linus Pauling Institute — Micronutrient Information Center — the authoritative scientific summary of nutrient biology including amino-acid metabolism
- NIH Office of Dietary Supplements — Fact Sheets Index — includes the official ODS sports-supplement statements on beta-alanine
- MedlinePlus — ALT (Alanine Aminotransferase) Blood Test
- MedlinePlus — Beta-Alanine
- ISSN Position Stand on Beta-Alanine (Trexler 2015) — consensus dose and safety statement from the International Society of Sports Nutrition
- PubMed — All research on alanine (~80,000+ papers)
- PubMed — Beta-alanine and carnosine research
Connections
- Alanine (Main Page)
- Alanine for Gluconeogenesis & Blood Sugar
- Beta-Alanine for Endurance
- Alanine for Immune Function
- Alanine for Liver Function (ALT)
- Beta-Alanine — the separate beta-isomer and carnosine precursor.
- All Amino Acids
- Glutamine
- Glycine
- Glutamic Acid
- Leucine
- Isoleucine
- Valine
- Histidine
- Taurine
- Arginine
- Vitamin B6 (ALT cofactor)
- Liver Disease
- Diabetes
- Blood Sugar
- Fasting
- Fatigue
- Lab Tests
- Eggs