Beta-Alanine for Endurance and High-Intensity Exercise
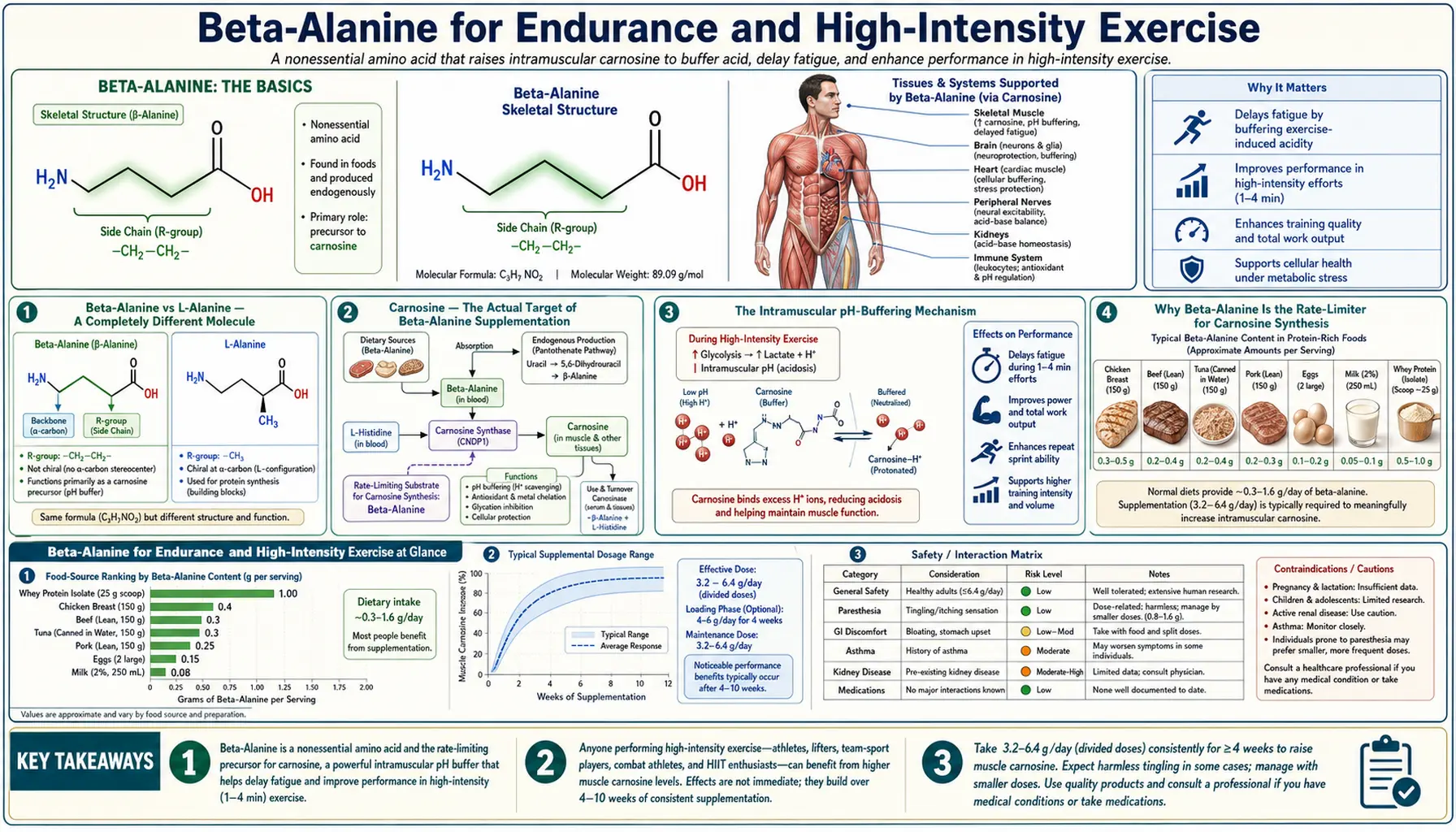
Beta-alanine is one of the most important misconceptions in sports nutrition: despite the shared name, it is biochemically and pharmacologically nothing like L-alanine. Beta-alanine (3-aminopropanoic acid) is a structural isomer in which the amino group sits on the beta-carbon rather than the alpha-carbon, and crucially, it is not incorporated into proteins. Its sole established role in human physiology is as the rate-limiting precursor for carnosine (beta-alanyl-L-histidine), a dipeptide present at high concentrations in skeletal muscle, where it functions as the principal intramuscular pH buffer during anaerobic exercise. Beta-alanine supplementation roughly doubles muscle carnosine concentration over 4-12 weeks of loading at 3.2-6.4 g/day — an effect achievable through no dietary route except concentrated meat consumption, and not achievable at all through L-alanine supplementation. Three decades of well-controlled randomized trials have established that beta-alanine produces a small but reliable improvement in exercise capacity for efforts in the 1-4 minute range (where intramuscular pH drops are most performance-limiting), making it one of only five supplements rated "A-tier" by the Australian Institute of Sport and the International Society of Sports Nutrition.
Table of Contents
- Beta-Alanine vs L-Alanine — A Completely Different Molecule
- Carnosine — The Actual Target of Beta-Alanine Supplementation
- The Intramuscular pH-Buffering Mechanism
- Why Beta-Alanine Is the Rate-Limiter for Carnosine Synthesis
- Dose and Loading Protocols (Harris / Hoffman)
- The Paraesthesia ("Tingle") Effect
- Documented Exercise-Capacity Effects
- Non-Responders and the Vegetarian Effect
- Other Putative Roles of Carnosine (Beyond Exercise)
- Stacking with Other Sports Supplements
- Cautions
- Key Research Papers
- Connections
- Featured Videos
Beta-Alanine vs L-Alanine — A Completely Different Molecule
The most important conceptual point about beta-alanine is that it is, despite the shared name, a fundamentally different molecule from L-alanine and serves a non-overlapping biological role. The two share only the molecular formula (C3H7NO2) and a stoichiometric similarity. Their biochemistry, transport, metabolism, and clinical applications are entirely separate.
The structural distinction is the position of the amino group relative to the carboxylic acid:
- L-alanine (alpha-alanine): The amino group is attached to the alpha-carbon — the carbon adjacent to the carboxylic acid group. This is the standard amino acid configuration shared by all 20 proteinogenic amino acids. L-alanine is incorporated into proteins via the standard ribosomal machinery and participates in transamination reactions catalyzed by alanine aminotransferase (ALT).
- Beta-alanine (3-aminopropanoic acid): The amino group is attached to the beta-carbon — one carbon further from the carboxylic acid. This single-bond difference makes beta-alanine non-proteinogenic. There is no transfer RNA that codes for beta-alanine, no aminoacyl-tRNA synthetase that recognizes it, and no ribosome that incorporates it into protein. It is also not a substrate for ALT (the enzyme is specific for the alpha-amino-acid stereochemistry).
The two amino acids are also transported differently. L-alanine uses the System A and System ASC sodium-dependent transporters (SNAT1, SNAT2, ASCT1, ASCT2). Beta-alanine uses a completely different transporter, the taurine/GABA transporter family (TauT/SLC6A6), which it shares with taurine and GABA. This is the biochemical reason that taurine and beta-alanine compete for uptake at the muscle cell membrane and at the blood-brain barrier — a fact relevant when stacking these two supplements.
The only metabolic connection between L-alanine and beta-alanine in human physiology is that beta-alanine can be slowly produced from L-aspartate through aspartate decarboxylation in the liver, and from cytosine and uracil (pyrimidine bases) through their catabolic pathway. These endogenous sources produce only modest amounts of beta-alanine — nowhere near enough to saturate muscle carnosine synthesis, which is why oral supplementation produces such striking carnosine increases.
Carnosine — The Actual Target of Beta-Alanine Supplementation
Carnosine (beta-alanyl-L-histidine) is a dipeptide of beta-alanine and L-histidine connected by a standard peptide bond. It was discovered in 1900 by the Russian biochemist V.S. Gulewitsch in beef extract (hence the name "carnosine" from carnis, Latin for flesh). It is present at high concentrations — 5-25 mmol/kg dry muscle — in skeletal muscle of all vertebrates, with the highest concentrations in the fast-twitch (Type II) fibers that are most dependent on anaerobic glycolysis.
The functional importance of carnosine in muscle was established through the work of Roger Harris and his colleagues in the early 2000s. Three properties make carnosine uniquely well-suited for its buffering role:
- Optimal pKa — the imidazole side chain of the histidine moiety has a pKa of approximately 6.83, very close to physiological intramuscular pH during heavy exercise (which can drop from 7.0 at rest to 6.4 during maximal anaerobic exertion). This is the ideal pKa range for a chemical buffer — the dipeptide accepts protons (H+) most efficiently exactly in the pH window where buffering matters most. Free histidine has a similar pKa but is less effective because of its non-restricted geometry.
- High intramuscular concentration — carnosine accumulates at concentrations of 10-30 mM in trained sprinters and elite anaerobic athletes, making it quantitatively the most important non-bicarbonate buffer in skeletal muscle. By comparison, intramuscular phosphate buffers are typically 5-10 mM and intramuscular protein buffers contribute another 20-30 mM equivalent capacity.
- Cytosolic localization — carnosine is freely dissolved in the muscle cell cytosol rather than confined to organelles. This puts it precisely where the protons accumulate during anaerobic glycolysis (lactate dehydrogenase produces NADH and protons together with lactate from pyruvate). The cytosol is the active site of buffering, not the mitochondria or the sarcoplasmic reticulum.
Carnosine concentrations vary substantially between individuals, fiber types, and species. Sprinters have higher concentrations than endurance athletes. Type IIx fibers (fastest fast-twitch) have higher concentrations than Type I (slow oxidative). Men have approximately 25% higher baseline concentrations than women. Vegetarians have about 50% lower baseline concentrations than omnivores (because meat is the only dietary source of preformed beta-alanine). Genetic polymorphisms in carnosine synthetase and in carnosinase (the degradative enzyme) explain some of the remaining individual variability.
The whole-body half-life of muscle carnosine is approximately 8-10 weeks, which means that loading takes about 4 weeks to reach near-steady state and washout after discontinuation takes a similar time. This relatively slow turnover is why beta-alanine is dosed continuously rather than only on training/competition days, and why the timing of beta-alanine intake within a day matters very little.
The Intramuscular pH-Buffering Mechanism
To understand why intramuscular carnosine matters for exercise performance, it helps to understand the proton-accumulation problem during anaerobic exercise.
During high-intensity exercise that exceeds the rate at which oxygen can be supplied and used by the mitochondria, muscle cells rely on anaerobic glycolysis — the conversion of glucose to lactate — for ATP production. Each glucose molecule that goes through anaerobic glycolysis yields 2 ATP plus 2 lactate plus 2 protons (H+). The lactate is not the cause of the "burn" or the fatigue — it is the protons. The lactate is shipped to the liver via the Cori cycle and is, if anything, a useful aerobic fuel for the heart and slow-twitch fibers. The protons are what drop intramuscular pH from 7.0 at rest toward 6.4 at exhaustion.
The pH drop has multiple performance-limiting effects:
- Inhibition of phosphofructokinase-1 (PFK-1) — the rate-limiting glycolytic enzyme is allosterically inhibited by low pH. As pH drops, glycolytic flux slows, ATP production falls, and the muscle cannot sustain force.
- Reduced calcium sensitivity of the contractile apparatus — troponin-C's calcium-binding affinity is pH-dependent. Lower pH reduces the calcium binding and thereby reduces the activation of the actin-myosin cross-bridge cycle for a given calcium release from the sarcoplasmic reticulum.
- Direct inhibition of the calcium-release channel (RyR1) — ryanodine receptor 1 activity is also pH-sensitive. Low cytosolic pH reduces the magnitude of each calcium release event from the SR.
- Increased excitatory neurotransmitter release from peripheral pain afferents — the "burn" itself acts as a CNS-mediated negative feedback signal that reduces voluntary effort.
Carnosine's role is to absorb protons before they can reach the concentrations that trigger these performance-limiting effects. Each mole of carnosine's imidazole group can buffer one mole of H+, so the total buffering capacity scales linearly with intramuscular concentration. Doubling muscle carnosine (the typical effect of beta-alanine loading) approximately doubles the carnosine contribution to total intramuscular buffering capacity, which is enough to delay the onset of severe acidosis by tens of seconds in the 1-4 minute exercise window.
Why Beta-Alanine Is the Rate-Limiter for Carnosine Synthesis
Carnosine is synthesized intramuscularly by carnosine synthase (ATPGD1, also known as CARNS1), which catalyzes the ATP-dependent condensation of free beta-alanine and free L-histidine. The reaction is:
beta-alanine + L-histidine + ATP → carnosine + AMP + PPi
The reaction depends on both substrates being available at sufficient concentration. The key insight from the Harris 2006 and Hill 2007 studies is that L-histidine is normally present in muscle at saturating concentrations, while beta-alanine is normally present at sub-saturating concentrations. This makes beta-alanine the rate-limiting substrate. Oral histidine loading does not raise muscle carnosine. Oral beta-alanine loading does.
The mathematical implication is straightforward: increasing the bioavailable beta-alanine pool increases the substrate for carnosine synthase, which proportionally increases the muscle carnosine concentration over the weeks-long timescale that the muscle pool requires to equilibrate. The kinetics of the equilibration follow a roughly exponential approach to a new steady-state ceiling. Hill 2007 showed that 4 weeks of loading at 4.8 g/day raised muscle carnosine by 58%, and 10 weeks of loading at the same dose raised it by 80% — the additional 6 weeks added much less benefit than the first 4 weeks, consistent with an asymptotic approach to a ceiling.
The diet contains some preformed carnosine and anserine (the methylated carnosine analog found at higher concentrations in poultry and fish), but the intestinal enzyme serum carnosinase (CN1, CNDP1) hydrolyzes essentially all dietary carnosine in the gut before it reaches systemic circulation. The net effect is that almost all the beta-alanine that reaches muscle for carnosine synthesis must come from supplemental free beta-alanine, not from dietary carnosine. This is why concentrated meat consumption raises muscle carnosine only modestly compared to direct beta-alanine supplementation.
Dose and Loading Protocols (Harris / Hoffman)
The canonical beta-alanine loading protocols are derived from the Harris 2006 and Hoffman 2008 dose-finding studies, codified in the 2015 ISSN Position Stand by Trexler and colleagues. The consensus recommendations are:
- Loading dose: 4-6 g/day for 4-12 weeks, divided into smaller doses of 0.8-1.6 g taken every 3-4 hours throughout the day. The division into smaller doses is important not for efficacy but to minimize paraesthesia (see below). The 4-week endpoint represents the minimum required to reach approximately 50-60% of the steady-state ceiling; 12 weeks reaches approximately 80-90%.
- Maintenance dose: Once loading is complete, 1.2-2.4 g/day is sufficient to maintain the elevated muscle carnosine concentration indefinitely. Discontinuation results in a slow washout with a half-life of approximately 6-9 weeks; muscle carnosine returns toward baseline over 12-18 weeks.
- Sustained-release formulations: Beta-alanine is available in slow-release tablets (Decombaz 2012) that release the dose over 1-2 hours rather than producing the peak plasma concentration of immediate-release. The slow-release formulations dramatically reduce paraesthesia and allow once-daily dosing of the full 4-6 g without the tingle effect.
- Timing within a day: Does not matter for efficacy. Carnosine is built up over weeks; the time-of-day a given dose is taken has no impact on the steady-state concentration. Athletes may prefer to take beta-alanine away from training to avoid paraesthesia during sessions, or with food to slow absorption and reduce paraesthesia.
- Cycling on and off: Not biochemically necessary. Long-term studies up to 24 months have shown sustained safety. Some athletes choose to cycle off during the off-season to reduce supplement cost, with the understanding that they will need 4-12 weeks of reloading before the next competitive season.
Doses above 6 g/day do not produce proportionally greater muscle carnosine elevation — the carnosine synthase reaction reaches saturation, and the additional beta-alanine is either excreted unchanged in urine or oxidized via the dihydropyrimidine dehydrogenase pathway. Doses above 10 g/day are wasted and dramatically increase paraesthesia without performance benefit.
The Paraesthesia ("Tingle") Effect
The most distinctive feature of beta-alanine supplementation is the harmless paraesthesia effect that occurs within 15-30 minutes of taking a bolus dose larger than approximately 800 mg. It is described variously as "pins and needles," "skin tingles," or a "tropical itch" sensation, typically felt in the face, neck, hands, and torso. It subsides within 30-60 minutes and produces no lasting effect. Athletes initially mistake it for an allergic reaction or a stimulant effect; it is neither.
The mechanism is the activation of the MrgprD receptor (Mas-related G-protein-coupled receptor member D), a recently discovered class of GPCR expressed on a subset of cutaneous C-fiber sensory neurons. Beta-alanine is one of the only known endogenous ligands of MrgprD, and high plasma beta-alanine concentrations transiently depolarize MrgprD-expressing neurons, producing the tingle sensation. The same MrgprD pathway is also involved in itch sensation, which is why some users describe the sensation as itchy rather than tingly.
Practical management of paraesthesia includes:
- Divided dosing — the original Harris/Hoffman 0.8-1.6 g doses every 3-4 hours stays below the paraesthesia threshold for most users.
- Slow-release tablets — the Decombaz formulation flattens the peak plasma concentration enough to eliminate paraesthesia even at 1.6 g per dose.
- Taking with food — gastric emptying delays absorption and reduces peak plasma concentration; a 1.6 g dose taken with a meal produces minimal paraesthesia for most users.
- Habituation — the paraesthesia intensity decreases over weeks of consistent use as users either upregulate MrgprD desensitization or simply adapt to the sensation.
The paraesthesia is harmless and reversible. No serious adverse events have been linked to it in long-term studies. The clinical significance is essentially zero — it is a marker of the dose having been absorbed, not a sign of toxicity. Users who find it intolerable can use slow-release formulations to eliminate it entirely.
Documented Exercise-Capacity Effects
The Saunders 2017 BJSM meta-analysis (40 studies, 70 individual exercise outcomes, n=1,461 participants) is the largest aggregation of beta-alanine exercise trials. The findings are remarkably consistent and define the efficacy window:
- Exercise duration of 0.5 to 10 minutes: Overall effect size of approximately +2.85% improvement in performance (small but reliable). The 95% confidence interval excludes zero.
- Exercise duration of 1 to 4 minutes (the "sweet spot"): Effect size of approximately +4-5% — the largest and most reliable effects are in this window. This corresponds to events like 400m and 800m running, 200m freestyle swimming, 1000m rowing, and 30-second to 4-minute Wingate or CrossFit-type efforts.
- Exercise duration below 30 seconds: No meaningful benefit. Very short maximal efforts are powered by ATP-PCr (creatine phosphate) rather than glycolysis, so intramuscular pH does not drop enough to be performance-limiting. Creatine is the appropriate supplement for this duration range, not beta-alanine.
- Exercise duration above 10 minutes: Diminishing benefit. Long-duration exercise is oxidative rather than glycolytic, so the intramuscular pH does not drop into the carnosine-buffering range. Beta-alanine has minimal value for marathon running, long-distance cycling, or other endurance events without high-intensity surges.
- Exercise involving repeated sprint bouts: Beta-alanine improves performance in the later sprints of a series. Single sprints show no benefit; repeated sprints with short rest intervals (CrossFit, soccer, hockey, basketball) show consistent benefit because the cumulative pH drop is in the carnosine-buffering range.
Real-world translation in elite athletes is modest but meaningful. A 4% improvement in a 2000m rowing time means about 9 seconds faster for an elite 6:00 erg time — the difference between qualifying for and missing the Olympic team. For recreational athletes, the practical benefit is "a few more reps" or "a few more seconds" in the late stages of glycolytically demanding sessions. Beta-alanine is not a transformative supplement — it does not make slow athletes fast — but it produces a small reliable edge in the specific exercise domain where carnosine buffering matters.
Non-Responders and the Vegetarian Effect
Roughly 10-15% of beta-alanine users in published trials show muscle carnosine increases below the population mean. Several mechanisms have been identified:
- Serum carnosinase (CNDP1) polymorphisms — high-activity CNDP1 variants hydrolyze plasma carnosine and beta-alanyl-histidine intermediates more rapidly, reducing the carnosine pool available for muscle uptake. The CNDP1 CTG haplotype is associated with reduced beta-alanine efficacy.
- Baseline carnosine concentration — subjects who start with high baseline carnosine show smaller percentage increases because they are closer to the carnosine synthase saturation ceiling. Trained sprinters and meat-eaters often show smaller increases than untrained subjects and vegetarians.
- Carnosine synthase variants — rare polymorphisms in ATPGD1/CARNS1 produce lower enzyme activity and reduce the carnosine response to beta-alanine loading.
Conversely, vegetarians are the population most likely to show large beta-alanine responses. Because vegetarian diets contain essentially no preformed carnosine and minimal beta-alanine (which is concentrated in muscle meat), vegetarians typically have 30-50% lower baseline muscle carnosine than omnivores. Beta-alanine supplementation in vegetarian athletes therefore produces larger relative increases and larger associated performance improvements. The 2007 Derave trial in trained sprinters showed substantial benefit; vegetarian subjects in subsequent trials have shown approximately 50% larger response magnitudes.
Other Putative Roles of Carnosine (Beyond Exercise)
Beyond its established role as a muscle pH buffer, carnosine has additional biological properties that are areas of active research, though clinical translations remain incomplete:
- Anti-glycation — carnosine is one of the most potent endogenous inhibitors of advanced glycation end-product (AGE) formation. It binds to reactive carbonyl groups (methylglyoxal, glyoxal, 4-hydroxynonenal) and quenches them before they can crosslink with proteins. This has generated interest in carnosine as an anti-aging compound and as a complementary therapy in diabetic complications, though clinical evidence remains preliminary.
- Antioxidant activity — carnosine scavenges singlet oxygen, hydroxyl radicals, and peroxyl radicals. It chelates copper, iron, and zinc, all of which can drive Fenton-type radical chemistry. The functional significance of this in vivo is debated.
- Heavy metal chelation — carnosine binds copper, zinc, and other transition metals. Some research has explored zinc-L-carnosine as a gastric mucosal protectant (used clinically in Japan as polaprezinc for gastric ulcer healing).
- Neuroprotection — carnosine and its methylated relative homocarnosine are present in the brain (homocarnosine in CSF reflects GABA metabolism). Some preclinical evidence suggests neuroprotection in models of cerebral ischemia, but clinical trials in stroke have been small and inconclusive.
- Wound healing — topical and oral carnosine have been studied in diabetic ulcer healing. Effects are modest but the safety profile is excellent.
For the well-established role of beta-alanine in exercise specifically, the evidence base is strong. The other potential applications remain promising but unproven at clinical scale. Athletes should not expect beta-alanine to be a magic anti-aging or neuroprotection supplement — the safe and demonstrated use case is exercise capacity in glycolytically demanding sport.
Stacking with Other Sports Supplements
Beta-alanine is one of the "A-tier" evidence-supported sports supplements alongside creatine monohydrate, caffeine, sodium bicarbonate, and nitrates (beetroot). All five have independent mechanisms and can be stacked productively:
- Beta-alanine + creatine monohydrate — the most popular stack, with strong rationale. Creatine extends the ATP-PCr system that powers very short maximal efforts (5-20 seconds); beta-alanine extends the glycolytic system that powers slightly longer efforts (30 seconds to 4 minutes). The two work in adjacent energy systems and produce additive benefits in CrossFit-style mixed-domain sessions. The 2006 Stout trial showed additive effects on physical working capacity.
- Beta-alanine + sodium bicarbonate — both work as pH buffers but at different sites. Beta-alanine (via carnosine) buffers intramuscularly; sodium bicarbonate (via bicarbonate) buffers extracellularly in the blood, drawing protons out of muscle into circulation via the proton-lactate cotransporter. The two have additive effects, but sodium bicarbonate carries a high gastrointestinal side-effect burden (acute diarrhea) that limits its real-world use.
- Beta-alanine + caffeine — non-overlapping mechanisms (caffeine is central-nervous-system focused on perceived exertion and motor unit recruitment). Stacking is safe and produces additive effects on high-intensity exercise.
- Beta-alanine + beetroot/nitrate — nitrate works by improving mitochondrial efficiency and lowering the oxygen cost of submaximal exercise. Different mechanism from beta-alanine's pH buffering. The two are additive in middle-distance events.
- Beta-alanine + taurine — competes for the same TauT transporter. Some authors have suggested that chronic high-dose beta-alanine could deplete muscle taurine, but the magnitude appears small and clinically inconsequential. Taurine supplementation alongside beta-alanine prevents any theoretical depletion concern.
For more on the broader sports nutrition landscape, see our pages on Leucine (for muscle protein synthesis) and Glutamine (for recovery and gut barrier function).
Cautions
- Paraesthesia is harmless but distracting — use slow-release formulations or divided dosing if the tingle interferes with quality of life or training focus.
- No documented long-term safety issues — the longest published trials extend to 24 months at 3.2-6.4 g/day with no adverse events beyond paraesthesia. Beta-alanine is rated as "safe" by the ISSN, the AIS, and the European Food Safety Authority.
- Theoretical taurine depletion — via TauT competition. Magnitude is small but vegetarian endurance athletes (who may also have lower baseline taurine) could consider co-supplementing taurine 1-2 g/day.
- Pregnancy and lactation — no studies have evaluated beta-alanine safety in pregnancy. Pregnant and breastfeeding women should avoid supplementation in the absence of safety data.
- Children and adolescents — beta-alanine is not recommended for athletes under 18 due to lack of pediatric safety data. The principal use case (high-intensity competitive sport) is adult-focused anyway.
- Banned substance concerns — beta-alanine is not on the WADA Prohibited List and is allowed in all competitive sports. It is a naturally occurring molecule and produces no positive doping test.
- Distinction from L-alanine for the consumer — do not confuse the two when shopping. L-alanine supplements (typically marketed for blood sugar or general amino acid support) do not raise muscle carnosine and do not produce the exercise capacity benefits of beta-alanine. The label should specifically say "beta-alanine" (sometimes written as "β-alanine").
- Stack interactions with stimulants — many pre-workout products combine beta-alanine, caffeine, and other stimulants. The combined paraesthesia plus stimulant arousal can feel intense and is sometimes mistakenly attributed to one ingredient alone. Knowing which sensation comes from which ingredient helps users titrate the stack to their tolerance.
Key Research Papers
- Harris RC, Tallon MJ, Dunnett M, et al. (2006). The absorption of orally supplied beta-alanine and its effect on muscle carnosine synthesis in human vastus lateralis. Amino Acids 30(3):279-289. — DOI: 10.1007/s00726-006-0299-9
- Hill CA, Harris RC, Kim HJ, et al. (2007). Influence of beta-alanine supplementation on skeletal muscle carnosine concentrations and high intensity cycling capacity. Amino Acids 32(2):225-233. — DOI: 10.1007/s00726-006-0364-4
- Hoffman J, Ratamess NA, Faigenbaum AD, et al. (2008). Short-duration beta-alanine supplementation increases training volume and reduces subjective feelings of fatigue. Nutrition Research 28(1):31-35. — DOI: 10.1016/j.nutres.2007.11.004
- Trexler ET, Smith-Ryan AE, Stout JR, et al. (2015). International Society of Sports Nutrition position stand: Beta-alanine. Journal of the International Society of Sports Nutrition 12:30. — DOI: 10.1186/s12970-015-0090-y
- Saunders B, Elliott-Sale K, Artioli GG, et al. (2017). Beta-alanine supplementation to improve exercise capacity and performance: a systematic review and meta-analysis. British Journal of Sports Medicine 51(8):658-669. — DOI: 10.1136/bjsports-2016-096396
- Boldyrev AA, Aldini G, Derave W (2013). Physiology and pathophysiology of carnosine. Physiological Reviews 93(4):1803-1845. — DOI: 10.1152/physrev.00039.2012
- Derave W, Ozdemir MS, Harris RC, et al. (2007). Beta-Alanine supplementation augments muscle carnosine content and attenuates fatigue during repeated isokinetic contraction bouts in trained sprinters. Journal of Applied Physiology 103(5):1736-1743. — DOI: 10.1152/japplphysiol.00795.2007
- Stout JR, Cramer JT, Mielke M, et al. (2006). Effects of twenty-eight days of beta-alanine and creatine monohydrate supplementation. Journal of Strength & Conditioning Research 20(4):928-931. — DOI: 10.1519/R-18525.1
- Decombaz J, Beaumont M, Vuichoud J, et al. (2012). Effect of slow-release beta-alanine tablets on absorption kinetics and paresthesia. Amino Acids 43(1):67-76. — DOI: 10.1007/s00726-011-1169-7
- Sale C, Saunders B, Harris RC (2010). Effect of beta-alanine supplementation on muscle carnosine concentrations and exercise performance. Amino Acids 39(2):321-333. — DOI: 10.1007/s00726-009-0443-4
- Liu Q, Sikand P, Ma C, et al. (2012). Mechanisms of itch evoked by beta-alanine. Journal of Neuroscience 32(42):14532-14537. — DOI: 10.1523/JNEUROSCI.3509-12.2012
- Stegen S, Bex T, Vervaet C, et al. (2014). Beta-alanine dose for maintaining moderately elevated muscle carnosine levels. Medicine & Science in Sports & Exercise 46(7):1426-1432. — DOI: 10.1249/MSS.0000000000000248
PubMed Topic Searches
- PubMed: Beta-alanine and muscle carnosine
- PubMed: Beta-alanine and exercise performance
- PubMed: Carnosine intramuscular pH buffering
- PubMed: Beta-alanine paraesthesia mechanism
- PubMed: Carnosinase and beta-alanine non-responders
- PubMed: Carnosine anti-glycation and antioxidant roles
Connections
- Alanine Overview
- Alanine Benefits Hub
- Alanine for Gluconeogenesis & Blood Sugar
- Alanine for Immune Function
- Alanine for Liver Function (ALT)
- Histidine (Carnosine Partner)
- Taurine (TauT Transporter Sibling)
- Leucine
- Isoleucine
- Valine
- Glutamine
- Creatine Monohydrate
- Athletic Performance
- Beef (Dietary Carnosine)
- Chicken (Dietary Carnosine)
- Fatigue
- All Amino Acids
- Beta-Alanine — the standalone page for the carnosine-precursor amino acid compared throughout this article.