Alanine for Gluconeogenesis and Blood Sugar
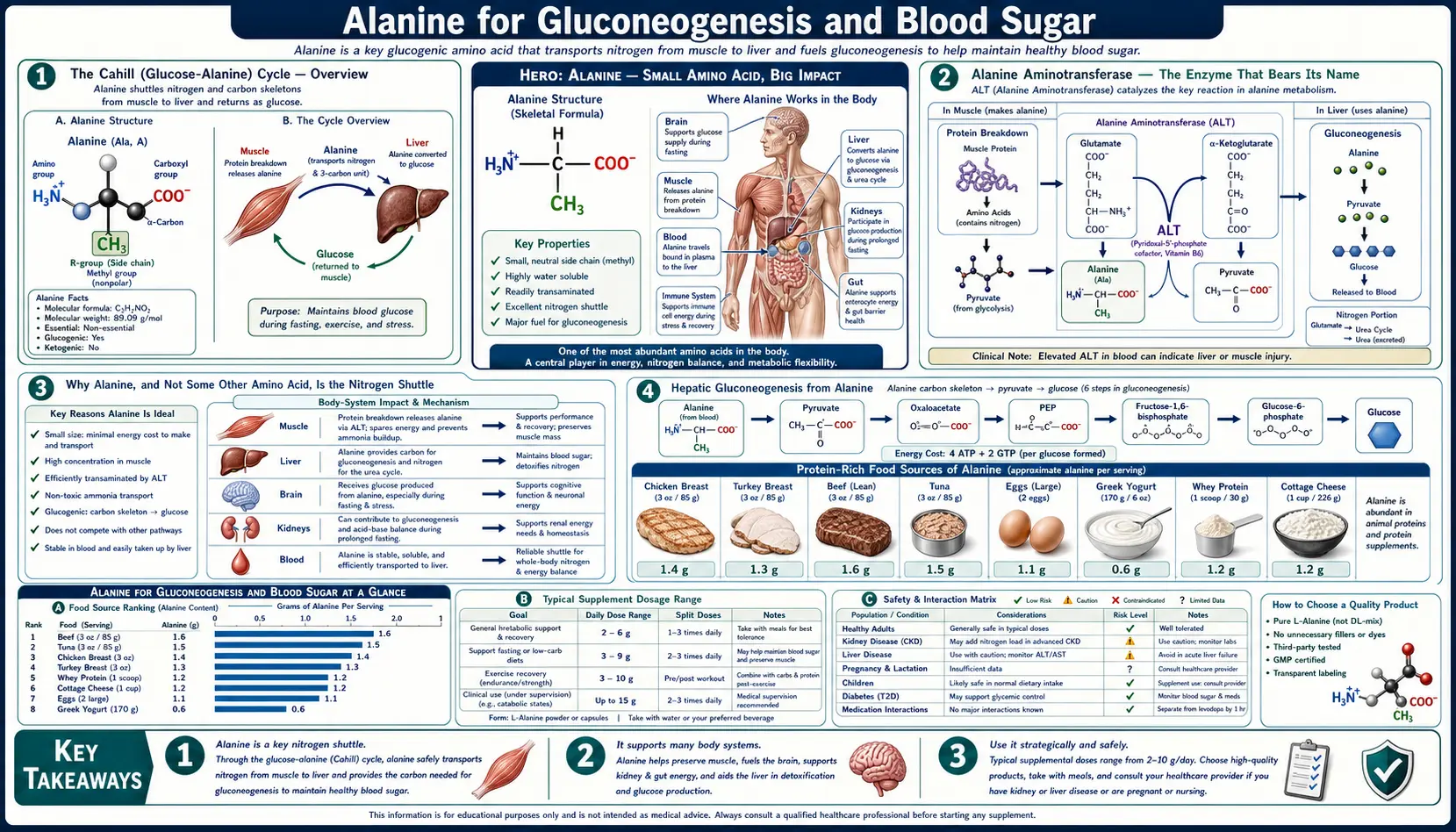
The glucose-alanine cycle — named the Cahill cycle after the Harvard physiologist George F. Cahill, Jr., whose 1970 series of Science and JCI papers established its quantitative importance — is the single most elegant inter-organ metabolic loop in human biochemistry. Skeletal muscle catabolizes branched-chain amino acids (leucine, isoleucine, valine) for local energy production during exercise and fasting, but the liberated nitrogen cannot be processed locally because muscle lacks a complete urea cycle. The nitrogen is therefore repackaged onto pyruvate by alanine aminotransferase (ALT) to make alanine, exported to the bloodstream, and disposed of safely by the hepatic urea cycle. In the same step, the liver harvests the pyruvate carbon skeleton for gluconeogenesis, returning glucose to the bloodstream to refuel the working muscle. Without this cycle, fasting, prolonged exercise, and protein turnover would each create an intolerable nitrogen-disposal crisis. This page walks through the biochemistry, the named ALT liver-panel enzyme that catalyzes the transamination, the physiology of overnight fasting and prolonged starvation, the 60-90% exercise-induced alanine flux, and alanine's direct role as a glucagon secretagogue.
Table of Contents
- The Cahill (Glucose-Alanine) Cycle — Overview
- Alanine Aminotransferase — The Enzyme That Bears Its Name
- Why Alanine, and Not Some Other Amino Acid, Is the Nitrogen Shuttle
- Hepatic Gluconeogenesis from Alanine
- Overnight Fasting and Prolonged Starvation
- Exercise-Induced Alanine Output (60-90% Increase)
- Alanine as a Glucagon Secretagogue
- Alanine, Insulin, and the Postprandial State
- Clinical Applications — Hypoglycemia, Hepatic Failure, Type 1 Diabetes
- Cautions
- Key Research Papers
- Connections
- Featured Videos
The Cahill (Glucose-Alanine) Cycle — Overview
The glucose-alanine cycle was first quantitatively characterized in human subjects by Philip Felig and George F. Cahill, Jr. at Harvard between 1968 and 1973. Earlier biochemists had identified the individual reactions, but Felig and Cahill's arteriovenous-difference studies in fasting men — measuring the concentration of alanine entering and leaving the leg via the femoral artery and vein — showed that alanine accounts for over half of all amino acid nitrogen released from skeletal muscle during fasting, and that the liver clears that alanine at a rate that quantitatively matches the rate of hepatic gluconeogenic glucose output. The cycle is now textbook biochemistry but was, at the time, a genuinely revolutionary unification of muscle physiology, hepatic biochemistry, and nitrogen disposal.
The cycle proceeds in a closed loop between skeletal muscle and liver:
- Glucose is delivered to muscle from the liver — either from hepatic glycogenolysis (the breakdown of stored glycogen) in the early hours of a fast, or from hepatic gluconeogenesis (de novo glucose synthesis) once glycogen stores begin to deplete around 8-12 hours of fasting.
- Muscle metabolizes glucose to pyruvate via glycolysis — the universal first 10-step pathway of glucose breakdown, ending at pyruvate. Pyruvate is the central junction between carbohydrate and amino-acid metabolism.
- Pyruvate accepts a nitrogen group from a branched-chain amino acid via ALT — alanine aminotransferase (ALT) catalyzes the reversible transfer of the amino group from glutamate to pyruvate, producing alanine and alpha-ketoglutarate. The nitrogen ultimately originates from branched-chain amino acids (leucine, isoleucine, valine) that have been transaminated by branched-chain aminotransferase (BCAT) onto alpha-ketoglutarate to form glutamate.
- Alanine is released into the bloodstream — muscle has a sodium-dependent alanine transporter that exports alanine into the venous drainage. Alanine concentrations in the femoral vein of a fasting subject exceed femoral artery concentrations by approximately 30-50%, the empirical signature of net muscle alanine release.
- Liver imports alanine via portal circulation — the liver expresses high levels of both alanine transporters and ALT. Hepatic ALT runs the same reaction in reverse, regenerating pyruvate and glutamate.
- Glutamate donates its amino group to the urea cycle — via glutamate dehydrogenase, glutamate releases its amino group as free ammonia, which enters carbamoyl phosphate synthetase I (CPS I) and is converted to urea for renal excretion. This is the safe-disposal step that muscle cannot perform locally.
- Pyruvate enters hepatic gluconeogenesis — pyruvate carboxylase converts pyruvate to oxaloacetate (requiring biotin as cofactor and ATP), and the rest of the gluconeogenic pathway (PEPCK, fructose-1,6-bisphosphatase, glucose-6-phosphatase) converts oxaloacetate back to glucose.
- Glucose is released into the bloodstream — closing the loop. The carbon skeleton has traveled from glucose to pyruvate to alanine to pyruvate and back to glucose, while one nitrogen has been moved from a branched-chain amino acid to urea.
The cycle achieves three goals simultaneously: continuous glucose delivery to glucose-dependent tissues (red blood cells, renal medulla, and to a substantial extent the brain), safe nitrogen disposal from peripheral tissues, and protein-sparing during prolonged fasting (because the liver can use alanine's carbon for glucose rather than catabolizing the same carbon from de novo amino-acid breakdown).
Alanine Aminotransferase — The Enzyme That Bears Its Name
Alanine aminotransferase (ALT, EC 2.6.1.2) is the enzyme that catalyzes the central transamination reaction of the glucose-alanine cycle:
L-alanine + alpha-ketoglutarate ↔ pyruvate + L-glutamate
The enzyme is reversible and uses pyridoxal-5-phosphate (the active form of vitamin B6) as an obligate cofactor. The reaction direction is determined by substrate concentrations and the cellular need: muscle ALT runs the reaction in the forward direction (consuming pyruvate and glutamate to produce alanine and alpha-ketoglutarate, supporting nitrogen export), while liver ALT runs it in reverse (consuming alanine to produce pyruvate, feeding gluconeogenesis and the urea cycle).
ALT has historical importance well beyond its biochemistry — it is by far the most-ordered biomarker on every standard liver panel ordered in clinical medicine worldwide. The historical name SGPT (serum glutamic-pyruvic transaminase) is still encountered in older literature and some non-US laboratories, but the modern designation ALT is now standard. Normal serum ALT in adults is typically below 30-40 U/L (some sources advocate even stricter cutoffs of 19 U/L for women and 30 U/L for men based on the 2002 Prati study). Concentrations of ALT inside hepatocytes are approximately 3,000-fold higher than serum levels, which is why even modest hepatocyte injury releases enough enzyme to produce measurable serum elevations.
The clinical interpretation of ALT elevation patterns is the gateway to all liver disease diagnosis — covered in detail on the companion Alanine and Liver Function page. For the gluconeogenesis story, the key point is that ALT is the limiting enzyme that allows the entire glucose-alanine cycle to function. Deficiency of pyridoxal-5-phosphate (vitamin B6) impairs ALT activity and can produce a mild functional impairment of nitrogen disposal — one of the lesser-known consequences of marginal B6 status.
Why Alanine, and Not Some Other Amino Acid, Is the Nitrogen Shuttle
A natural question is why alanine, specifically, evolved to carry nitrogen from periphery to liver. Several biochemical and physiological factors converge:
- Pyruvate is universally available. Glycolysis runs in every cell that uses glucose, which is essentially every cell. The carbon skeleton precursor for alanine (pyruvate) therefore costs nothing — it is sitting at the end of glycolysis waiting to be either reduced to lactate (anaerobic fate), oxidized to acetyl-CoA (aerobic fate), or transaminated to alanine (nitrogen-shuttle fate). The same is not true for the carbon skeletons of other potential shuttle molecules.
- Alanine is non-toxic at high concentrations. Plasma alanine in a fasting individual can reach 400-500 micromolar without any toxic effect. Compare this to free ammonia, which is severely neurotoxic above 100 micromolar. The whole point of the shuttle is to move nitrogen safely — alanine's low toxicity is what makes high-flux nitrogen transport tolerable.
- Alanine is structurally simple and metabolically cheap. Of all the amino acids, only glycine is simpler. Alanine's small size means it costs minimal ATP to synthesize (just one transamination from already-existing pyruvate) and minimal energy to transport across cell membranes.
- Hepatic uptake of alanine is rapid. The liver has high-affinity alanine transporters (System A) on the sinusoidal membrane and runs ALT at high flux. The first-pass clearance of portal alanine is approximately 70-80% in a fasting subject, so very little alanine "escapes" the liver back into systemic circulation.
- The reverse direction is metabolically useful. The liver can immediately use the pyruvate output of hepatic ALT for gluconeogenesis. This couples nitrogen disposal to a useful synthetic output (glucose) rather than requiring a separate disposal pathway for the carbon skeleton.
Glutamine is the secondary nitrogen shuttle in human metabolism, carrying nitrogen primarily from muscle to gut and from gut to liver via a slightly different route. Together, alanine and glutamine account for over 70% of inter-organ amino acid flux in the fasting state. Alanine dominates muscle-to-liver flow; glutamine dominates muscle-to-gut and gut-to-liver flow.
Hepatic Gluconeogenesis from Alanine
Once alanine reaches the liver, the carbon skeleton can be diverted to gluconeogenesis. The pathway from alanine to glucose has six key enzymatic steps:
- ALT (alanine aminotransferase) converts alanine + alpha-ketoglutarate to pyruvate + glutamate. This is the entry step.
- Pyruvate carboxylase (a biotin-dependent enzyme located in the mitochondrial matrix) converts pyruvate + CO2 + ATP to oxaloacetate. This is a key regulatory step — acetyl-CoA activates pyruvate carboxylase, coupling fatty acid oxidation to gluconeogenesis.
- PEPCK (phosphoenolpyruvate carboxykinase) converts oxaloacetate + GTP to phosphoenolpyruvate (PEP) + CO2 + GDP. The PEPCK gene is acutely induced by glucagon and chronically induced by glucocorticoids — this is the major hormonally regulated step.
- The reverse of glycolysis converts PEP back up through fructose-1,6-bisphosphate, with one key irreversible step: fructose-1,6-bisphosphatase hydrolyzes fructose-1,6-bisphosphate to fructose-6-phosphate. This step is allosterically inhibited by fructose-2,6-bisphosphate and by AMP, and reciprocally activates the opposing glycolytic enzyme phosphofructokinase-1 (PFK-1).
- Fructose-6-phosphate is converted to glucose-6-phosphate by the reversible enzyme phosphohexose isomerase.
- Glucose-6-phosphatase hydrolyzes glucose-6-phosphate to free glucose, which exits the hepatocyte via GLUT2 into the bloodstream. This is the final irreversible step and the most liver-specific enzyme of the pathway (it is also expressed in renal cortex but absent from muscle and adipose).
The overall energetics are unfavorable — producing one molecule of glucose from two molecules of alanine costs 4 ATP and 2 GTP (the cost of pyruvate carboxylase, PEPCK, and several phosphorylation steps). This energy is supplied by hepatic fatty acid oxidation, which is why fasting people break down fat to provide the energy that allows the liver to make glucose from amino acids.
The fraction of total hepatic glucose output coming from alanine depends on the duration of fasting. In the postprandial state, hepatic glucose output is essentially zero (the liver is taking up glucose). After 8-12 hours of fasting, glycogenolysis is the dominant source. After 24-48 hours, glycogen is largely depleted and gluconeogenesis takes over — with alanine providing roughly 40-50% of the gluconeogenic substrate, lactate providing 30-40%, and glycerol (from triglyceride breakdown) providing the remainder. By 72 hours of fasting, alanine is still the largest single amino acid contributor, but the brain has started to adapt to ketone bodies and overall glucose demand has decreased.
Overnight Fasting and Prolonged Starvation
The glucose-alanine cycle is the physiological basis of how humans tolerate overnight fasting (and longer fasts) without becoming hypoglycemic. The cycle operates continuously between meals but ramps up sharply during the post-absorptive state (4-12 hours after a meal) and dominates by the early-fasting state (12-24 hours):
- Hours 0-4 post-meal (absorptive state): Dietary glucose is being absorbed and circulating insulin is high. Hepatic glucose output is suppressed. Muscle is taking up glucose for glycogen synthesis. Plasma alanine is roughly 250-350 micromolar (highest postprandially, reflecting dietary amino acid absorption).
- Hours 4-12 post-meal (post-absorptive state): Insulin falls, glucagon rises. Hepatic glycogenolysis becomes the dominant source of glucose. Muscle alanine release begins to increase. Plasma alanine remains stable around 250-300 micromolar.
- Hours 12-24 (early fasting): Hepatic glycogen is approaching depletion (a 70-kg adult has roughly 75-100 g hepatic glycogen, providing about 6 hours of basal glucose output). Gluconeogenesis is ramping up. Alanine release from muscle increases approximately 30-50% over baseline. The transition from glycogenolysis-dominant to gluconeogenesis-dominant occurs around hour 18-24.
- Days 1-3 (early starvation): Gluconeogenesis is fully dominant. Alanine is the largest single substrate. Plasma alanine actually falls at this point to roughly 150-200 micromolar — the liver is clearing it faster than muscle can supply it. This decrease in plasma alanine has been proposed by Felig as a rate-limiting step in further gluconeogenesis, partially explaining the protein-sparing adaptation that begins around day 3-5 of fasting.
- Days 5-21 (sustained starvation): Hepatic ketogenesis is fully active. The brain adapts to use beta-hydroxybutyrate for roughly 70% of its energy needs by day 14. Glucose demand falls. Alanine release from muscle decreases (protein-sparing). Net nitrogen loss in urine falls from ~12 g/day to ~3-5 g/day, all of this adaptation reducing the demand on the alanine shuttle.
The implication is that the glucose-alanine cycle is most metabolically stressed not during prolonged starvation (where the body has adapted) but during the early fasting transition (12-72 hours), and most importantly during exercise overlaid on a fasting state (such as fasted morning training in athletes). Pyridoxal-5-phosphate (B6) cofactor availability becomes a meaningful constraint on ALT activity during these high-flux states.
Exercise-Induced Alanine Output (60-90% Increase)
Felig and Wahren's 1971 leg-exercise studies in human subjects demonstrated quantitatively the role of alanine in supporting hepatic glucose output during sustained exercise. The findings have been replicated in dozens of subsequent studies and are now textbook physiology:
- Femoral arteriovenous (A-V) alanine difference in resting fasting subjects is approximately 30-50 micromolar (the leg releases alanine into the venous drainage).
- During moderate-intensity leg exercise (cycling at ~50-60% VO2max), the alanine A-V difference increases 60-90% above baseline. The absolute alanine output per minute from one leg roughly doubles.
- Splanchnic alanine uptake increases proportionally — the liver clears the additional alanine load and converts it to glucose at increased rates, exactly matching the muscle's increased glucose demand.
- The cycle scales with exercise intensity up to ~75% VO2max. Above that threshold, alanine flux plateaus and lactate flux begins to dominate (the lactate-alanine shift reflects the increasing fraction of anaerobic glycolysis at high intensity).
- The cycle scales with exercise duration. During exercise lasting longer than 60 minutes, alanine flux remains elevated and contributes increasing fractions of total hepatic gluconeogenic substrate as glycogen depletes.
The practical implication for endurance athletes is that adequate dietary protein intake is metabolically important not just for muscle synthesis (the obvious story) but for maintaining the substrate pool that supports the glucose-alanine cycle during long training sessions. Branched-chain amino acid availability becomes the actual rate-limiting input — the BCAA nitrogen is what gets transaminated to alanine. This is one mechanistic explanation for the older observation that BCAA supplementation modestly improves endurance performance in glycogen-depleted states.
It is worth noting that the glucose-alanine cycle is distinct from (and complementary to) the Cori cycle, which moves lactate from muscle to liver and glucose from liver to muscle. The Cori cycle handles the anaerobic carbon load; the glucose-alanine cycle handles the nitrogen load. Both run simultaneously during exercise.
Alanine as a Glucagon Secretagogue
Alanine has a remarkable second metabolic role that closes a feedback loop in the glucose-alanine cycle: it directly stimulates glucagon secretion from pancreatic alpha cells. Glucagon is the hormone that activates hepatic gluconeogenesis — so a rising plasma alanine concentration (the substrate for gluconeogenesis) directly recruits the hormonal signal that triggers gluconeogenesis (glucagon). This is one of the most elegant substrate-hormone feedback loops in metabolism.
The mechanism involves alanine uptake by alpha cells through a sodium-dependent transporter, which depolarizes the cell, activates voltage-gated calcium channels, and triggers glucagon vesicle exocytosis. The effect is dose-dependent and acutely measurable — intravenous alanine infusion in healthy subjects raises plasma glucagon within minutes. This is the basis for the clinical "alanine load test" that has been used in research to assess alpha cell function and to characterize patients with glucagonoma.
The same loop also explains a clinically important observation: a high-protein meal raises plasma glucagon as well as plasma insulin. Insulin and glucagon normally oppose each other (insulin promotes anabolism, glucagon promotes catabolism), but after a protein meal, both rise simultaneously. The insulin is needed to anabolize the absorbed amino acids into muscle protein; the glucagon is needed to mobilize hepatic glucose to prevent the insulin from causing hypoglycemia in the absence of dietary carbohydrate. The relative ratio of the two hormones is fine-tuned by the proportion of carbohydrate to protein in the meal.
This dual hormonal response is why pure protein meals (e.g., a chicken breast and a salad) do not cause the same blood glucose excursion that an equivalent carbohydrate meal would — the simultaneous glucagon response counterbalances the insulin response. People with type 1 diabetes lack the insulin response but still mount a glucagon response, which is part of why protein meals can produce gradual late post-meal hyperglycemia in type 1 diabetes that requires extended insulin coverage.
Alanine, Insulin, and the Postprandial State
After a mixed meal containing both carbohydrate and protein, several metabolic events involving alanine occur in rapid succession:
- Dietary carbohydrate is absorbed and raises plasma glucose, which triggers insulin secretion.
- Dietary protein is digested to free amino acids and small peptides, absorbed across the intestinal epithelium, and delivered to the liver via the portal vein. Alanine is among the most abundant amino acids in dietary protein.
- The liver clears approximately 70% of absorbed alanine on the first pass — some goes into hepatic protein synthesis, some goes into immediate gluconeogenesis (which is suppressed but not zero in the postprandial state), and some goes into glutamate via reverse ALT for further metabolic distribution.
- The amino acids that escape first-pass hepatic clearance (mostly the branched-chain amino acids, which the liver clears poorly) reach systemic circulation and provide the substrate for muscle protein synthesis under the anabolic drive of insulin.
- Plasma alanine concentration rises by ~30-50% above the fasting baseline within 2 hours of a meal and returns toward baseline by 4-6 hours.
The postprandial alanine rise is one of the metabolic signals that contributes to the "second meal effect" — the well-documented observation that a high-protein meal in the morning improves glucose tolerance at lunchtime. The mechanism is partially through increased alanine availability for hepatic gluconeogenesis during the late post-absorptive phase of the morning meal, smoothing the glucose curve and preventing the rebound hypoglycemia that would otherwise occur and trigger a counter-regulatory hyperglycemia at lunch.
This is also why protein-fortified breakfasts (eggs, Greek yogurt, cottage cheese, smoked salmon) tend to produce more stable mid-morning blood sugar than carbohydrate-only breakfasts (pastries, cereals) even in non-diabetic subjects. The increased alanine pool from breakfast protein supports a smoother gluconeogenic transition into the late morning. For more on dietary protein strategies for blood sugar regulation, see our Blood Sugar Remedies page.
Clinical Applications — Hypoglycemia, Hepatic Failure, Type 1 Diabetes
The glucose-alanine cycle becomes clinically relevant in several specific conditions:
- Reactive hypoglycemia — the phenomenon of post-meal blood glucose drops below 70 mg/dL, typically 2-4 hours after a high-carbohydrate meal. Pure carbohydrate meals trigger a large insulin response without the simultaneous glucagon counter-regulation that a protein-containing meal would provide. The result is a rapid glucose clearance followed by a glycemic "valley" if hepatic gluconeogenesis cannot ramp up quickly enough. Adding protein (with its accompanying alanine) to every meal blunts the insulin response, provides simultaneous glucagon counter-regulation, and supplies gluconeogenic substrate — all of which prevent the reactive hypoglycemia.
- Hepatic failure — when liver function is severely impaired (cirrhosis, fulminant hepatic failure), the glucose-alanine cycle collapses. Hepatic gluconeogenesis fails and patients become prone to spontaneous fasting hypoglycemia. Alanine accumulates in plasma because the liver cannot clear it (plasma alanine in advanced cirrhosis can reach 800-1000 micromolar versus normal 250-350). The accumulated alanine cannot be processed and the BCAA-to-AAA (aromatic amino acid) ratio falls, contributing to hepatic encephalopathy. This is the rationale behind BCAA-enriched parenteral and enteral nutrition formulations for advanced liver disease — they support the muscle's ability to maintain the BCAA-derived nitrogen flow to alanine without overloading the failing liver.
- Type 1 diabetes — type 1 diabetics lack endogenous insulin but retain normal alpha-cell glucagon responses. The glucagon response to dietary protein continues unopposed by insulin, producing a delayed hyperglycemia from a protein-containing meal that becomes visible 3-5 hours after eating. Modern insulin pump algorithms account for this by extending bolus insulin coverage based on protein content, not just carbohydrate content. Plasma alanine elevations are typical in poorly controlled type 1 diabetes.
- Inborn errors of gluconeogenesis — rare genetic deficiencies of pyruvate carboxylase, PEPCK, fructose-1,6-bisphosphatase, or glucose-6-phosphatase (von Gierke disease, glycogen storage disease type 1) all impair the ability to convert alanine to glucose and produce severe fasting hypoglycemia, lactic acidosis, and hyperalaninemia. These children require frequent small meals and continuous overnight nasogastric feeding to prevent fasting hypoglycemia.
- Severe protein-energy malnutrition (kwashiorkor, marasmus) — chronic dietary protein deficiency depletes muscle BCAA pools, which depletes the alanine shuttle, which impairs the ability to maintain fasting blood glucose. Affected children have a paradoxical fatty liver despite caloric deprivation, partially because impaired alanine availability blocks gluconeogenesis and reroutes carbon to hepatic triglyceride synthesis.
Cautions
- Chronic kidney disease (CKD) — impaired urea excretion can produce mild rises in plasma amino acids including alanine. Patients with advanced CKD should consult their nephrologist before taking any amino-acid supplement. Renal-failure-specific amino acid formulations are typically essential-amino-acid-enriched and minimize non-essential amino acid loading.
- Hepatic encephalopathy — in advanced cirrhosis, supplemental amino acids (especially aromatic amino acids and methionine) can worsen encephalopathy. BCAA-enriched formulations are preferred. Routine alanine supplementation is not indicated.
- Pyruvate carboxylase deficiency and other rare congenital gluconeogenic defects — alanine cannot be converted to glucose in these children, and alanine loading worsens lactic acidosis. These are rare but warrant mention.
- Inborn errors of urea cycle — carbamoyl phosphate synthetase deficiency, ornithine transcarbamylase deficiency, and similar conditions impair the disposal of nitrogen from alanine. High-protein meals trigger acute hyperammonemia in these patients. Protein restriction is the standard management.
- Drug-induced ALT elevation — many medications can elevate ALT (statins, acetaminophen, isoniazid, methotrexate, herbal products including kava and green tea extract at high doses, anabolic steroids). Elevated ALT in this setting reflects hepatocyte injury, not an alanine problem — the alanine cycle itself continues to function but is now being assessed against an injured liver background.
- Plasma alanine elevation in clinical interpretation — should always be interpreted alongside other plasma amino acid concentrations. Isolated mild plasma alanine elevation is usually a metabolic snapshot rather than a pathology. Persistent hyperalaninemia (above 500 micromolar) in the fasting state warrants metabolic workup for hepatic dysfunction or inborn errors.
Key Research Papers
- Felig P, Pozefsky T, Marliss E, Cahill GF (1970). Alanine: key role in gluconeogenesis. Science 167(3920):1003-1004. — DOI: 10.1126/science.167.3920.1003
- Felig P (1973). The glucose-alanine cycle. Metabolism 22(2):179-207. — PubMed
- Felig P, Marliss E, Pozefsky T, Cahill GF (1970). Amino acid metabolism in the regulation of gluconeogenesis in man. American Journal of Clinical Nutrition 23(7):986-992. — PubMed
- Cahill GF (2006). Fuel metabolism in starvation. Annual Review of Nutrition 26:1-22. — DOI: 10.1146/annurev.nutr.26.061505.111258
- Wahren J, Felig P, Ahlborg G, Jorfeldt L (1971). Glucose metabolism during leg exercise in man. JCI 50(12):2715-2725. — DOI: 10.1172/JCI106683
- Ahlborg G, Felig P, Hagenfeldt L, Hendler R, Wahren J (1974). Substrate turnover during prolonged exercise in man. JCI 53(4):1080-1090. — DOI: 10.1172/JCI107889
- Owen OE, Felig P, Morgan AP, Wahren J, Cahill GF (1969). Liver and kidney metabolism during prolonged starvation. JCI 48(3):574-583. — DOI: 10.1172/JCI106013
- Garber AJ, Karl IE, Kipnis DM (1976). Alanine and glutamine synthesis and release from skeletal muscle. Journal of Biological Chemistry 251(3):826-835. — PubMed
- Rothman DL, Magnusson I, Katz LD, Shulman RG, Shulman GI (1991). Quantitation of hepatic glycogenolysis and gluconeogenesis in fasting humans (NMR). Science 254(5031):573-576. — DOI: 10.1126/science.1948033
- Genuth SM, Castro J (1974). Effect of oral alanine administration in fasting obese subjects. Metabolism 23(4):375-386. — PubMed
- Snell K (1980). Muscle alanine synthesis and hepatic gluconeogenesis. Biochemical Society Transactions 8(2):205-213. — PubMed
- Petersen KF, Dufour S, Befroy D, Garcia R, Shulman GI (2004). Impaired mitochondrial activity in the insulin-resistant offspring of patients with type 2 diabetes. NEJM 350(7):664-671. — DOI: 10.1056/NEJMoa031314
- Nuttall FQ, Schweim KJ, Gannon MC (2006). Effect of orally administered alanine on plasma glucose, insulin and glucagon concentrations. British Journal of Nutrition 95(2):358-365. — DOI: 10.1079/BJN20051622
- Hagenfeldt L, Wahren J (1971). Human forearm muscle metabolism during exercise. Scandinavian Journal of Clinical and Laboratory Investigation 28(4):359-369. — PubMed
PubMed Topic Searches
- PubMed: Glucose-alanine (Cahill) cycle
- PubMed: Alanine aminotransferase (ALT) biochemistry
- PubMed: Alanine during exercise and nitrogen shuttle
- PubMed: Alanine and glucagon secretion
- PubMed: Fasting gluconeogenesis substrates
- PubMed: Urea cycle and amino acid nitrogen
Connections
- Alanine Overview
- Alanine Benefits Hub
- Beta-Alanine for Endurance
- Alanine for Immune Function
- Alanine for Liver Function (ALT)
- Glutamine (Secondary Nitrogen Shuttle)
- Leucine (BCAA Nitrogen Donor)
- Isoleucine
- Valine
- Glutamic Acid (Transamination Partner)
- Vitamin B6 (ALT Cofactor)
- Blood Sugar Remedies
- Fasting
- Diabetes
- Liver Disease
- Fatigue
- Lab Tests
- All Amino Acids