Boron for Bone Density
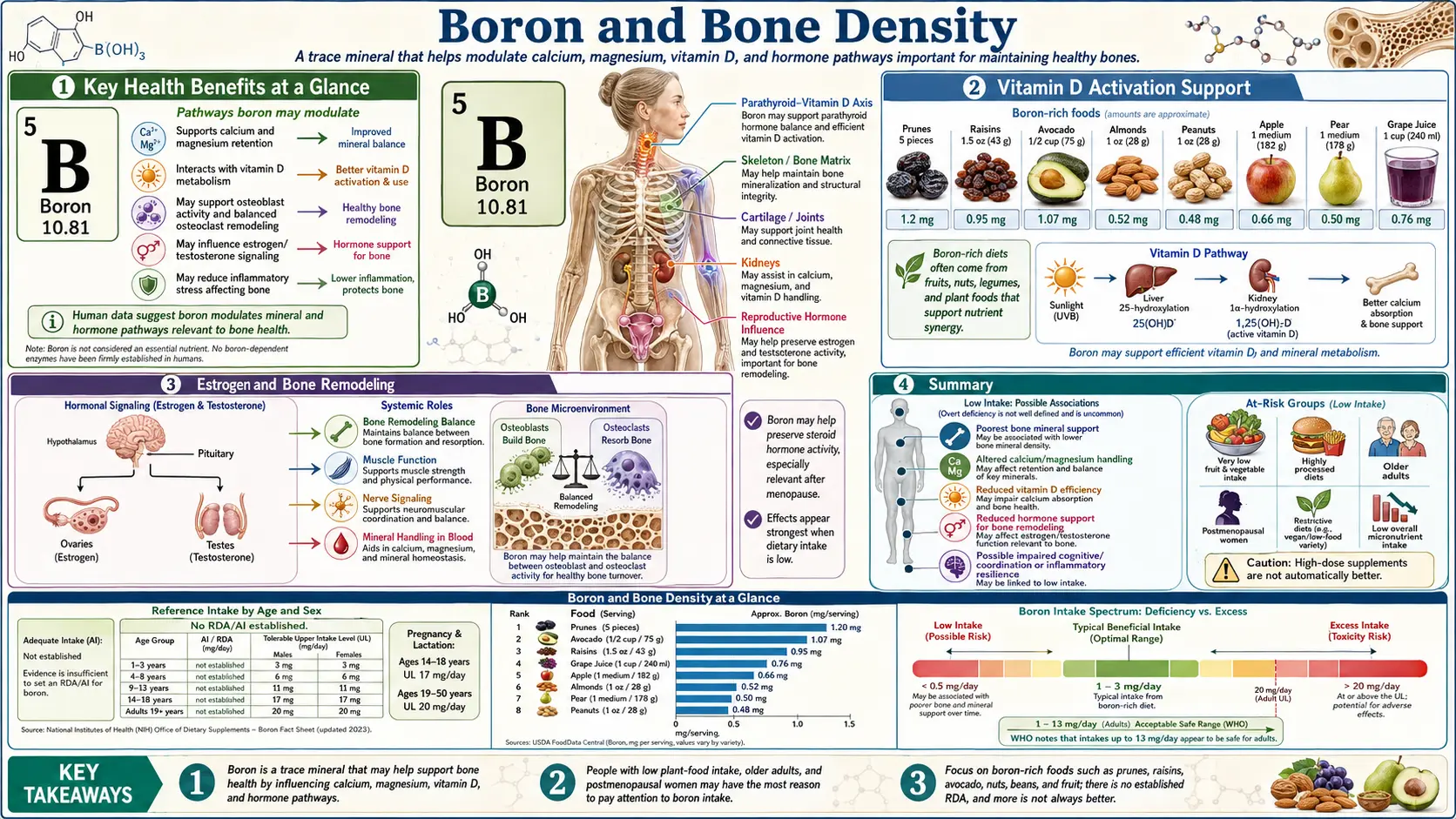
Boron is the ultratrace mineral that turned out to be unexpectedly central to skeletal biology. The total body store is only a few milligrams, but dietary intake exerts outsized leverage on calcium retention, magnesium utilization, vitamin D activation, and steroid-hormone metabolism — four of the most important determinants of bone mineral density. Forrest Nielsen's pioneering USDA metabolic-ward studies in the 1980s and 1990s showed that boron deprivation (0.25 mg/day) impaired mineral balance and depressed circulating estradiol and testosterone, while modest repletion (3 mg/day) reversed those effects within weeks. This deep-dive walks through the mechanisms, the clinical evidence in postmenopausal women and aging men, and the practical implications for anyone working to prevent or reverse osteoporosis.
Table of Contents
- Why Boron Matters to Bone Despite Being a Trace Mineral
- Calcium Metabolism and Urinary Calcium Loss
- Vitamin D Activation and Half-Life Extension
- Magnesium Retention and Bone Matrix Enzymes
- Estrogen, Testosterone, and Bone Remodeling
- Osteoblast Stimulation and Osteoclast Suppression
- Bone Turnover Markers and Hydroxyapatite Quality
- Osteoporosis Prevention and Postmenopausal Bone Loss
- The Nielsen USDA Metabolic Ward Studies
- Geographic Epidemiology — Soil Boron and Bone Disease
- Practical Dosing, Forms, and Safety
- Cautions and Drug Interactions
- Key Research Papers
- Connections
- Featured Videos
Why Boron Matters to Bone Despite Being a Trace Mineral
The total body content of boron in an adult human is approximately 3–20 mg — roughly the amount in a single small pinch of borax powder. By comparison, the body contains about 1.2 kg of calcium and 25 g of magnesium. On a mass basis, boron looks unimportant. But on a functional basis, boron operates as a metabolic regulator: it does not contribute structural mass to bone, it shapes how the body handles the minerals and hormones that do.
This is the same pattern seen with other ultratrace minerals like selenium (which makes up <15 mg in the body but is required for every selenoprotein), iodine (essential for thyroid hormone synthesis at <50 mg total), and chromium (proposed glucose regulator at trace levels). Boron joins this category of regulatory micronutrients whose clinical significance vastly exceeds their mass.
The biochemistry that explains this is boron's avidity for cis-hydroxyl groups (-OH groups on adjacent carbons of a sugar or steroid ring). Boric acid, the form in which boron exists in body fluids, forms reversible covalent complexes with these cis-diols. This single chemical preference connects boron to:
- Sugar-containing biological molecules — ribose, NAD+, S-adenosylmethionine (SAMe), nucleotides, glycoproteins, and proteoglycans of cartilage all carry cis-diol groups that can bind boron and have their activity modulated
- Steroid hormones — the hydroxylation patterns of cortisol, estradiol, testosterone, and 1,25(OH)2D3 all contain potential boron-binding sites that influence transport, half-life, and receptor engagement
- Phospholipid cell membranes — the inositol phospholipids that anchor membrane signaling proteins contain cis-diols that interact with boric acid
- Hyaluronic acid and the proteoglycans of cartilage and bone matrix — the glycosaminoglycan backbones are rich in cis-diol groups
The convergence of these binding partners is why boron supplementation produces effects across the bone-matrix, hormone, and cartilage systems simultaneously rather than affecting a single isolated pathway.
Calcium Metabolism and Urinary Calcium Loss
The single most reproducible boron-on-bone effect documented in controlled human trials is a reduction in urinary calcium excretion. The original observation came from Forrest Nielsen's USDA metabolic ward studies. After acclimating postmenopausal women to a low-boron diet (0.25 mg/day) for 119 days, urinary calcium loss was elevated. When boron was restored to 3 mg/day, urinary calcium dropped by approximately 44% — a magnitude that translates directly to improved calcium balance.
The mechanism is multifactorial:
- Vitamin D-mediated intestinal absorption — boron extends the half-life of 25(OH)D and supports its conversion to 1,25(OH)2D3, which upregulates the duodenal calbindin-D9k and TRPV6 calcium channels that drive transcellular calcium uptake. More calcium absorbed means less compensatory bone resorption needed
- Renal calcium reabsorption — boron-supported magnesium retention (see below) supports the magnesium-dependent components of the renal calcium reabsorptive machinery in the distal convoluted tubule and the thick ascending limb of the loop of Henle
- Reduced parathyroid hormone drive — when serum calcium is maintained, parathyroid hormone (PTH) does not rise compensatorily, and the PTH-driven bone resorption cascade is not activated
- Estradiol-mediated effects — boron-elevated estradiol independently suppresses osteoclast bone resorption, further reducing the calcium that gets liberated from bone and then either reincorporated or excreted
The clinical significance: net daily calcium balance is the difference between intake-absorption (positive) and urinary plus fecal losses (negative). In postmenopausal women, the average bone-mineral-density loss of 1–2%/year represents roughly 200–400 mg of bone calcium lost into urine annually. A 44% reduction in urinary calcium loss, sustained over decades, plausibly accounts for a meaningful slowing of postmenopausal bone density decline.
Vitamin D Activation and Half-Life Extension
The vitamin D activation cascade involves two sequential hydroxylations: hepatic 25-hydroxylase (CYP2R1) converts cholecalciferol to 25-hydroxyvitamin D, then renal 1-alpha-hydroxylase (CYP27B1) converts 25(OH)D to the hormonally active 1,25-dihydroxyvitamin D (calcitriol). Boron influences both steps.
The mechanism likely involves boron's interaction with the cytochrome P450 enzyme family. Both CYP2R1 and CYP27B1 are P450 enzymes whose activity is sensitive to the redox state of the heme iron and the geometry of the substrate-binding pocket. Boron may modulate either parameter through its general effect on steroid-hydroxylation chemistry.
Two clinically relevant downstream effects:
- Extended 25(OH)D half-life — Nielsen's metabolic ward studies showed approximately 20% higher serum 25(OH)D concentrations in boron-replete subjects on identical dietary vitamin D intake. The boron-treated subjects had more of their existing vitamin D stores available for renal activation
- Higher 1,25(OH)2D3 production — the active hormone, which drives the calbindin-D and TRPV6 expression that controls intestinal calcium absorption and the osteoblast differentiation that drives bone formation
The seasonal implication: cutaneous vitamin D synthesis falls to near zero at latitudes above 40 degrees during the winter months. Maintaining adequate 25(OH)D through the winter depends entirely on body stores, dietary intake, and supplement use. Boron's effect on extending 25(OH)D half-life could be particularly valuable during these months, when every unit of available vitamin D needs to be conserved.
For more on vitamin D in bone health, see our Vitamin D3 page.
Magnesium Retention and Bone Matrix Enzymes
Approximately 60% of total body magnesium is stored in bone — partly as part of the hydroxyapatite crystal lattice itself (where Mg2+ can substitute for Ca2+) and partly bound to the bone matrix as a reservoir that can be mobilized to maintain serum magnesium during dietary deficiency. Magnesium is required as an enzymatic cofactor for alkaline phosphatase (the enzyme that catalyzes the dephosphorylation reactions central to bone mineralization), for proper PTH secretion and end-organ responsiveness, and for the ATP-dependent calcium transport pumps in bone cells.
Boron supplementation reduces urinary magnesium excretion in a fashion parallel to its calcium-sparing effect. Nielsen's studies documented decreased urinary magnesium loss with boron repletion, particularly in subjects on baseline marginal magnesium intake. The mechanism likely involves boron-mediated effects on the renal magnesium reabsorption machinery (TRPM6 channels in the distal convoluted tubule), although the exact molecular interaction is not fully mapped.
The downstream consequences of better magnesium retention for bone:
- Maintained alkaline phosphatase activity — the bone-specific isoform is required for the dephosphorylation steps that allow hydroxyapatite crystal nucleation. Without adequate magnesium, alkaline phosphatase function is impaired and bone mineralization slows even when calcium and phosphate are abundant
- Hydroxyapatite crystal quality — magnesium influences the size and stability of hydroxyapatite crystals. Magnesium-poor bone tends to form excessively large, brittle crystals that compromise the elastic-modulus characteristics of cortical bone. Magnesium-replete bone has smaller, more uniform crystals with better mechanical properties
- Normal PTH responsiveness — magnesium-deficient patients have functional hypoparathyroidism (the parathyroid gland fails to secrete PTH normally) and end-organ resistance to PTH. Boron-supported magnesium adequacy preserves the PTH axis that maintains calcium-phosphorus homeostasis
- Vitamin D activation — both hepatic 25-hydroxylase and renal 1-alpha-hydroxylase require magnesium as a cofactor. Boron's magnesium-sparing effect therefore amplifies its direct effect on vitamin D activation
For more on magnesium specifically, see our Magnesium page and Magnesium Replenishment.
Estrogen, Testosterone, and Bone Remodeling
The estradiol decline at menopause is the dominant driver of postmenopausal osteoporosis. Estradiol normally restrains osteoclast bone resorption through multiple mechanisms: it directly suppresses osteoclast differentiation from monocyte precursors, it accelerates osteoclast apoptosis, and it shifts the RANKL/osteoprotegerin (OPG) ratio in osteoblasts toward more OPG (which sequesters RANKL and prevents it from activating osteoclasts). Loss of estradiol at menopause therefore unleashes osteoclast activity, and bone resorption accelerates relative to bone formation.
Boron's most striking hormonal effect in postmenopausal women is partial restoration of circulating estradiol. Nielsen's studies documented that 3 mg/day boron in postmenopausal women elevated serum 17-beta-estradiol from baseline depleted levels by approximately two-fold on average. The mechanism appears to be a combination of:
- Reduced hepatic clearance — boron may slow the cytochrome P450-mediated hydroxylation reactions that convert estradiol to less-active 2-hydroxy and 16-hydroxy metabolites
- Aromatase support — aromatase (CYP19A1) converts androstenedione to estrone in adipose and bone tissue. In postmenopausal women, this is the dominant source of circulating estradiol (ovarian production has ceased), and boron may support aromatase activity
- SHBG modulation — sex hormone-binding globulin sequesters circulating estradiol and testosterone in inactive form. Lower SHBG means more bioactive free hormone. Boron has been documented to reduce SHBG in supplementation trials
The implications for bone are direct: higher bioactive estradiol means less osteoclast activity, less bone resorption, and slower bone mineral density decline. This is the boron-on-bone mechanism that most clearly applies to postmenopausal women specifically, distinct from the calcium and vitamin D effects that apply more universally.
For men, the parallel boron-on-bone effect is via testosterone. The Naghii 2011 study (8 men, 10 mg/day boron for one week) documented a 28% increase in free testosterone, a 39% reduction in SHBG, and reductions in inflammatory markers (CRP, TNF-alpha, IL-6). For aging men with declining testosterone and increasing fracture risk, this hormone-protective effect contributes to boron's bone-protective profile. See Boron for Hormone Balance for the detailed hormone deep-dive.
Osteoblast Stimulation and Osteoclast Suppression
Bone is a dynamic tissue, continuously remodeled by the coordinated action of bone-forming osteoblasts and bone-resorbing osteoclasts. Bone mineral density at any given age is the integrated balance of these opposing processes. Boron acts on both sides of the equation, and on both at favorable angles.
Osteoblast effects — In vitro studies of cultured human osteoblasts show that boron at physiological concentrations (10–100 ng/mL boric acid):
- Increases osteoblast proliferation rate
- Increases alkaline phosphatase expression (the bone-mineralization-marker enzyme)
- Increases osteocalcin secretion (the vitamin K-dependent osteoblast protein that binds calcium and incorporates into bone matrix)
- Increases type I collagen synthesis (the dominant bone-matrix protein)
- Increases RUNX2 expression (the master transcription factor for osteoblast differentiation)
Osteoclast effects — The same in vitro work shows that boron suppresses osteoclast differentiation from RANK-expressing monocyte precursors. The mechanism is partly direct (boron may interfere with the cytoskeletal rearrangements required for osteoclast ruffled-border formation) and partly indirect through estradiol elevation (estradiol independently inhibits osteoclastogenesis through OPG upregulation).
The animal model evidence confirms that these in vitro effects translate to whole-bone outcomes. Boron-supplemented chicks have greater cortical bone thickness, higher trabecular bone volume, and improved mechanical strength on three-point bending tests. Ovariectomized rats (a standard animal model of postmenopausal osteoporosis) show partial restoration of bone density with boron supplementation, mimicking what the Nielsen human studies show for biochemical markers.
Bone Turnover Markers and Hydroxyapatite Quality
Modern bone-disease research uses serum and urinary biochemical markers to assess the rate of bone formation and resorption in real time, complementing the slower dual-energy X-ray absorptiometry (DXA) measurement of bone mineral density. Boron supplementation has been associated with favorable shifts in these markers:
- Bone formation markers UP
- Osteocalcin (the carboxylated, vitamin-K-dependent protein produced by mature osteoblasts and incorporated into bone matrix)
- Procollagen type I N-terminal propeptide (P1NP) — the cleavage fragment released as type I collagen is synthesized
- Bone-specific alkaline phosphatase (the osteoblast-derived isoform of total alkaline phosphatase)
- Bone resorption markers DOWN
- Urinary deoxypyridinoline (DPD) — the cross-linking molecule of mature type I collagen released into urine when osteoclasts resorb bone
- C-terminal telopeptide (CTX) and N-terminal telopeptide (NTX) of type I collagen — released into circulation by osteoclast-mediated bone resorption
- Tartrate-resistant acid phosphatase 5b (TRAP-5b) — the osteoclast-derived enzyme
The combination of increased formation markers and decreased resorption markers indicates a favorable shift in the remodeling balance — the same direction produced by bisphosphonates and SERMs (selective estrogen receptor modulators) used as pharmaceutical osteoporosis treatments, although the magnitude of effect is generally smaller with boron supplementation.
Hydroxyapatite crystal quality. Beyond the quantitative bone-density measurement, bone quality (the elastic-modulus, fracture-toughness, and fatigue-resistance properties) depends on hydroxyapatite crystal size and orientation, on collagen cross-linking maturity, and on the mineral-matrix ratio. Boron-supported magnesium retention favors smaller, more uniform hydroxyapatite crystals. Boron-supported alkaline phosphatase activity favors more uniform mineralization. Boron-mediated effects on collagen synthesis favor better matrix structure. The composite effect on bone quality may be more clinically important than the modest effect on DXA-measured density alone — bone quality determines fracture risk independently of bone mineral density.
Osteoporosis Prevention and Postmenopausal Bone Loss
Osteoporosis affects approximately 200 million women worldwide and is responsible for over 8.9 million fractures annually. Hip fracture in particular carries a 20% one-year mortality and a 50% rate of permanent loss of independent function in those who survive. Preventing osteoporosis is therefore one of the most consequential nutritional interventions in aging adults.
The conventional preventive regimen centers on adequate calcium (1000–1200 mg/day from diet plus supplements), vitamin D (800–2000 IU/day), weight-bearing exercise, and avoidance of bone-toxic habits (smoking, excessive alcohol, chronic glucocorticoid use). Boron is increasingly recognized as a fourth nutritional pillar, alongside calcium, vitamin D, and magnesium — the convergence of its calcium-sparing, vitamin D-extending, magnesium-retaining, and estradiol-restoring effects make it uniquely multifaceted for postmenopausal bone protection.
The boron-on-osteoporosis evidence comes from three converging directions:
- Geographic epidemiology — in regions with higher soil boron and dietary intake (Turkey, parts of Israel, certain wine-producing regions of California and Italy), osteoporosis and osteoarthritis incidence are lower. In regions with low soil boron (Jamaica, parts of New Zealand, the Mauritius islands), arthritis rates are dramatically higher
- Controlled human metabolic studies — Nielsen's USDA work and subsequent confirmatory trials documenting improved mineral balance and elevated estradiol with 3 mg/day boron supplementation in postmenopausal women
- Animal model trials — ovariectomized rats and chicks showing boron-induced increases in cortical thickness, trabecular bone volume, and mechanical strength
The integrated implication: postmenopausal women interested in preserving bone mineral density should consider boron supplementation at 3–6 mg/day as a low-cost, low-risk adjunct to standard calcium/vitamin D/magnesium/vitamin K2 supplementation and weight-bearing exercise. The intervention does not replace bisphosphonate or SERM therapy in established osteoporosis, but it complements them and may be sufficient as primary prevention in osteopenic women without yet-established disease.
For comprehensive postmenopausal bone management, see also our Osteoporosis page, Calcium and Bone Health, and Manganese and Bone Formation.
The Nielsen USDA Metabolic Ward Studies
The foundational human evidence for boron-on-bone comes from a series of metabolic ward studies conducted by Forrest H. Nielsen at the USDA Grand Forks Human Nutrition Research Center beginning in 1981 and continuing through the 1990s. The study design was rigorous: subjects (typically postmenopausal women aged 48–82) were housed in a metabolic research unit for prolonged periods, fed precisely controlled diets, and had their urine and stool collected for complete mineral balance calculations. This level of dietary control is unique to metabolic ward research and provides causality evidence that observational epidemiology cannot.
The seminal Nielsen 1987 paper (FASEB Journal 1:394) reported on 12 postmenopausal women fed a low-boron diet (0.25 mg/day) for 119 days followed by 3 mg/day supplementation for 48 days. The key findings:
- Urinary calcium loss decreased by approximately 44% with boron supplementation
- Urinary magnesium loss decreased by approximately 33%
- Serum 17-beta-estradiol approximately doubled (from depleted baseline to near-premenopausal levels)
- Serum testosterone roughly doubled
- The effects were more pronounced in subjects who were simultaneously magnesium-deficient (suggesting a synergy between boron and magnesium)
Subsequent Nielsen papers confirmed and extended these findings in different populations (premenopausal women, men, athletes) and at different dose levels. The 3 mg/day dose has emerged as the modal "benefit" dose in the literature, with 6–10 mg/day showing additional incremental effect in some studies but with diminishing returns above that.
The historical significance of the Nielsen studies cannot be overstated: they took boron from being considered a nutritionally inert curiosity to being recognized as a probably-essential trace mineral with measurable effects on mineral metabolism and steroid hormone biology. The fact that the changes were demonstrated at dietary intake levels achievable through normal food consumption (3 mg/day is the boron content of approximately one large avocado plus a handful of almonds) made the results immediately clinically relevant rather than purely a curiosity of pharmacologic dosing.
Geographic Epidemiology — Soil Boron and Bone Disease
Epidemiologic studies across multiple continents have documented a strikingly consistent inverse correlation between regional boron intake (driven primarily by soil boron content, which determines the boron content of locally grown produce and drinking water) and the prevalence of arthritis and osteoporosis.
Rex Newnham, an Australian biochemist working in the 1960s and 1970s, was the first to systematically document this pattern. His observations:
- Mauritius and Jamaica — low soil boron, dietary intake typically <1 mg/day, arthritis prevalence in adults over 50: 50–70%
- United States and Western Europe — moderate soil boron, dietary intake 1–2 mg/day, arthritis prevalence: 20–30%
- Carnarvon (Western Australia) — high boron from local geology, dietary intake 3–6 mg/day, arthritis prevalence: <1%
- Israel — high boron in tap water (5–15 mg/L in some regions), arthritis prevalence approximately 0.4%
The pattern is replicated for hip-fracture incidence and osteoporosis prevalence in independent studies. While epidemiologic correlations do not prove causation (cultural diet, exercise patterns, sun exposure for vitamin D, and many other variables differ between these regions), the consistency of the inverse boron-bone-disease correlation across populations as diverse as Caribbean islanders, Middle Eastern populations, and remote Australian communities makes random confounding implausible.
The follow-up controlled trial by Newnham himself: a double-blind, placebo-controlled study of 6 mg/day boron supplementation in osteoarthritis patients reported that 50% of the boron group experienced clinically meaningful symptom improvement, compared to 10% of placebo. See Boron for Arthritis & Joints for the detailed joint-disease deep-dive.
Practical Dosing, Forms, and Safety
Typical dietary intake in modern Western populations is 0.5–1.5 mg/day, well below the 3 mg/day documented as supporting optimal mineral balance in Nielsen's metabolic ward studies. This shortfall reflects the depletion of soil boron in modern agriculture (boron is consumed by crops and not replaced by typical fertilization programs) and the displacement of boron-rich whole foods by processed alternatives in modern diets.
Boron-rich foods (mg boron per 100 g):
- Raisins: 4.5 mg
- Dried apricots: 2.1 mg
- Avocado: 2.1 mg
- Almonds: 2.8 mg
- Brazil nuts: 1.7 mg
- Hazelnuts: 2.5 mg
- Prunes: 1.9 mg
- Lentils: 0.7 mg
- Beans (kidney, navy, lima): 0.5–1.0 mg
- Apples, pears, grapes: 0.3–0.5 mg
A diet that includes 50–100 g/day of mixed nuts, dried fruit, and avocado will naturally provide 3–5 mg of boron without supplementation. Where this is not practical, supplementation is the realistic alternative.
Supplement forms — the available oral supplements are:
- Boron citrate — the most common form, well absorbed, typically dosed at 3–6 mg elemental boron per capsule
- Boron glycinate — chelated form, similarly well absorbed
- Boron aspartate — chelated form
- Calcium fructoborate — a naturally occurring complex of boron with calcium and fructose, originally identified in plant tissue, marketed specifically for joint health (studied in arthritis trials at 1.5–6 mg/day)
- Sodium tetraborate (borax) — the cheapest source, used historically and still used by some self-experimenters at low dose (¼ teaspoon in 1 L water, providing ~125 mg per liter, drunk in small daily portions). Not a typical supplement-industry preparation but technically equivalent
The form does not appear to matter much for general supplementation — boron is well absorbed (>85% absorption from typical doses regardless of form) and circulates as boric acid. For joint-specific indications, calcium fructoborate has the most trial data and is reasonable to prefer.
Dose — the standard supplementation range for general bone-protective effects is 3 mg/day, taken with food. Higher doses up to 6–10 mg/day are reasonable for postmenopausal women interested in maximizing the estradiol effect or for men interested in the testosterone effect. The Institute of Medicine has set the Tolerable Upper Intake Level at 20 mg/day for adults; this is the dose at which observable adverse effects begin to occur in chronic studies, and there is a generous safety margin between the effective dose (3 mg) and the upper limit (20 mg).
Cautions and Drug Interactions
- Pregnancy — high-dose boron (>10 mg/day) is theoretically a concern in pregnancy based on animal teratogenicity studies at very high doses. Normal dietary intake (1–3 mg/day) is safe and is in fact required for normal embryonic development. The cautious approach is to avoid supraphysiologic supplementation in pregnancy and rely on dietary sources
- Acute toxicity — the LD50 for boric acid in adults is approximately 5–20 g (single ingestion). This is far above any conceivable supplement dose. Symptoms of acute toxicity at very high doses include nausea, vomiting, diarrhea, skin rash, lethargy, and (rarely) seizures
- Chronic toxicity — sustained intake of >100 mg/day for months has been associated with weight loss, hair loss, and dermatologic complaints. Supplementation at 3–10 mg/day has no documented chronic toxicity in any clinical trial
- Estrogen-sensitive conditions — women with estrogen-receptor-positive breast cancer history, endometriosis, or uterine fibroids should discuss boron supplementation with their oncologist or gynecologist. The estradiol-elevating effect, while modest, is real and may not be desirable in these contexts
- Hyperparathyroidism — in primary hyperparathyroidism, supplementation strategies should be coordinated with the treating endocrinologist. Boron's calcium-sparing effect could theoretically exacerbate hypercalcemia in this population
- Kidney disease — boron is renally excreted. In chronic kidney disease (GFR <30), boron accumulation could theoretically occur with chronic supplementation. Doses should be conservative or avoided in advanced CKD
- Drug interactions — minimal documented interactions. Boron does not significantly affect cytochrome P450-mediated drug metabolism at typical supplement doses
Key Research Papers
- Nielsen FH, Hunt CD, Mullen LM, Hunt JR (1987). Effect of dietary boron on mineral, estrogen, and testosterone metabolism in postmenopausal women. FASEB Journal. — PubMed
- Nielsen FH (2014). Update on human health effects of boron. Journal of Trace Elements in Medicine and Biology. — PubMed
- Pizzorno L (2015). Nothing boring about boron. Integrative Medicine (Encinitas). — PubMed
- Naghii MR et al. (2011). Comparative effects of daily and weekly boron supplementation on plasma steroid hormones and proinflammatory cytokines. Journal of Trace Elements in Medicine and Biology. — PubMed
- Newnham RE (1994). Essentiality of boron for healthy bones and joints. Environmental Health Perspectives. — PubMed
- Hunt CD (1996). Biochemical effects of physiological amounts of dietary boron. Journal of Trace Elements in Experimental Medicine. — PubMed
- Hakki SS et al. (2010). Boron regulates mineralized tissue-associated proteins in osteoblasts. Journal of Trace Elements in Medicine and Biology. — PubMed
- Devirian TA, Volpe SL (2003). The physiological effects of dietary boron. Critical Reviews in Food Science and Nutrition. — PubMed
- Meacham SL et al. (1995). Effect of boron supplementation on blood and urinary calcium, magnesium, and phosphorus, and urinary boron in athletic and sedentary women. American Journal of Clinical Nutrition. — PubMed
- Scorei R, Mitrut P, Petrisor I, Scorei I (2011). A double-blind, placebo-controlled pilot study to evaluate the effect of calcium fructoborate on systemic inflammation and dyslipidemia markers for middle-aged subjects with hypercholesterolemia. Biological Trace Element Research. — PubMed
- Palacios C (2006). The role of nutrients in bone health, from A to Z. Critical Reviews in Food Science and Nutrition. — PubMed
- Penland JG (1994). Dietary boron, brain function, and cognitive performance. Environmental Health Perspectives. — PubMed
PubMed Topic Searches
- PubMed: Boron and bone mineral density (postmenopausal)
- PubMed: Boron and calcium/magnesium balance
- PubMed: Boron and vitamin D activation
- PubMed: Boron, estradiol, testosterone, SHBG
- PubMed: Boron, osteoblast and osteoclast effects
- PubMed: Calcium fructoborate trials
Connections
- Boron Overview
- Boron Benefits Hub
- Boron for Hormone Balance
- Boron for Cognitive Function
- Boron for Arthritis & Joints
- Osteoporosis
- Arthritis
- Calcium
- Magnesium
- Silicon
- Phosphorus
- Manganese
- Calcium and Bone Health
- Manganese and Bone Formation
- Vitamin D3
- Vitamin K2
- Testosterone
- DHEA-S
- Collagen
- Magnesium Replenishment
- All Minerals