Threonine for Collagen and Connective Tissue
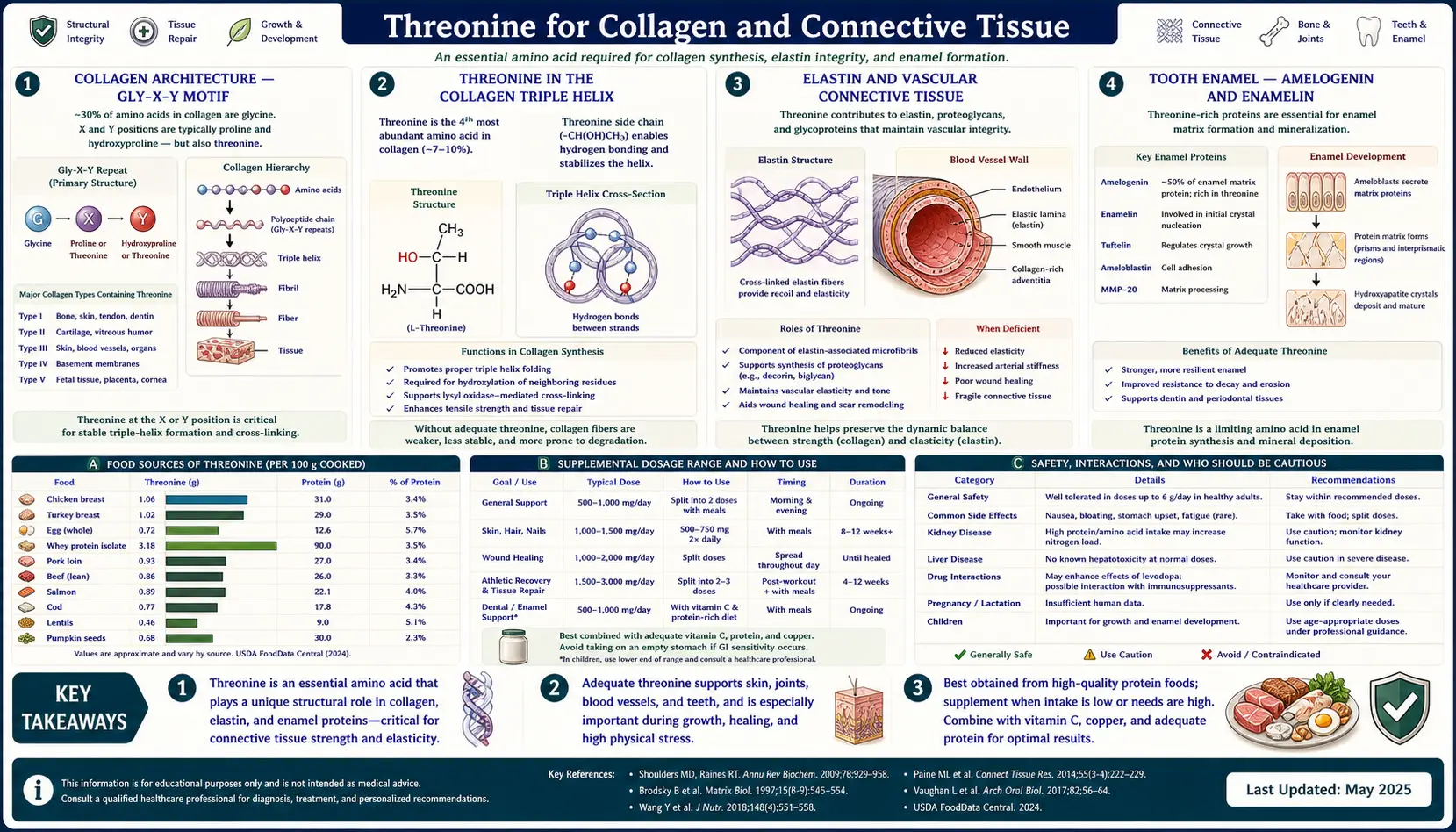
Threonine is the quiet partner in the collagen story. Glycine and proline get the headlines — correctly — because together they make up about 50% of every collagen triple helix and are the residues responsible for the iconic Gly-X-Y repeat that defines the helix geometry. But threonine has a different and equally essential role: it occupies the X and Y positions in many of the most cross-linked sequences, it appears as a phosphorylation site in collagen-regulating kinases, and most distinctively it dominates the tooth-enamel proteins amelogenin and enamelin, where it is the most abundant amino acid residue in the molecule. This page examines where threonine sits in the connective-tissue protein family, why scurvy-like collagen failure can occur from threonine inadequacy even with adequate vitamin C, and how dental, vascular, and dermal connective tissue all depend on it.
Table of Contents
- Collagen Architecture — The Gly-X-Y Motif
- Threonine in the Collagen Triple Helix
- Elastin and Vascular Connective Tissue
- Tooth Enamel — Amelogenin and Enamelin
- Bone Matrix Proteins
- Cartilage and Joint Proteoglycans
- Wound Healing and Connective Tissue Repair
- Crosslink Chemistry and Stability
- Aging Connective Tissue
- Clinical Supplementation
- Cautions
- Key Research Papers
- Connections
- Featured Videos
Collagen Architecture — The Gly-X-Y Motif
Collagen is the single most abundant protein in the human body, accounting for approximately 30% of all body protein by mass. There are at least 28 distinct collagen subtypes, but the structural feature shared by all of them is the triple helix — three polypeptide chains wound around each other in a tight right-handed superhelix, stabilized by hydrogen bonds and by post-translationally modified hydroxyproline and hydroxylysine residues.
The defining sequence motif of collagen is the Gly-X-Y repeat, where every third amino acid is glycine, and the X and Y positions are occupied by a variety of other amino acids, most commonly proline at X and 4-hydroxyproline at Y. Glycine is mandatory at every third position because its tiny side chain (just a hydrogen atom) is the only residue small enough to fit at the center of the triple helix where the three chains converge. Any substitution of glycine for a larger amino acid at this position kinks the helix and produces severe connective-tissue disease — osteogenesis imperfecta is the textbook example, caused by single-amino-acid substitutions of glycine in type I collagen.
The X and Y positions face outward from the helix axis and tolerate considerable amino acid diversity. About 25-30% of X positions are proline and about 25% of Y positions are 4-hydroxyproline. The remaining 40-50% of X and Y positions are filled by other amino acids including threonine, serine, lysine, glutamate, alanine, and others. The exact sequence at these positions determines collagen subtype, helix stability, post-translational modification pattern, and the binding interfaces for integrins, glycoproteins, and other extracellular matrix components.
Threonine in the Collagen Triple Helix
Threonine occupies approximately 2-3% of residues in mature type I collagen (the major collagen of skin, bone, and tendon), making it less abundant than glycine, proline, alanine, glutamate, or arginine, but considerably more abundant than tryptophan (which is absent from collagen entirely), histidine, or methionine. In type II collagen (the major collagen of cartilage), threonine content is closer to 3-4%. In type III collagen (reticular fibers, blood vessels, gut wall), threonine content is comparable to type I.
The role of threonine in collagen is twofold:
- Structural contribution to specific telopeptide regions. The N-terminal and C-terminal telopeptides of collagen molecules (the short non-helical extensions at each end of the procollagen) contain threonine residues that participate in the inter-molecular crosslinks that bind collagen molecules into the mature fibril. These crosslinks are formed by the enzyme lysyl oxidase acting on lysine and hydroxylysine residues, and the surrounding amino acid context — including threonine — determines the geometry and stability of the resulting crosslink.
- Substrate for O-glycosylation in certain collagen types. Collagens that contain galactose-O-hydroxylysine modifications also occasionally carry threonine-linked O-glycans in specific subtypes, particularly collagen XV and XVIII (the so-called "multiplexin" collagens found in basement membranes). These O-glycan modifications affect the interaction of collagen with other extracellular matrix components.
Because threonine is essential and cannot be synthesized de novo, inadequate dietary threonine creates a hard ceiling on collagen synthesis rate, just as inadequate vitamin C creates a ceiling on hydroxyproline formation. The clinical presentation of severe threonine deficiency in animals includes impaired wound healing, weak skin and tendons, and gut barrier failure — phenotypically similar to scurvy in some respects, though the molecular mechanism is different.
For more on the collagen-building partner amino acids, see our pages on Glycine, Proline, and Lysine, all of which work together with threonine in the collagen synthesis pathway.
Elastin and Vascular Connective Tissue
Elastin is the second major fibrous protein of connective tissue and is the dominant protein responsible for the elastic recoil of arteries, lungs, skin, and the bladder. Unlike collagen, elastin is not assembled into a precise triple helix — instead, the elastin precursor tropoelastin is secreted into the extracellular space where it self-aggregates and is crosslinked by lysyl oxidase into rubber-like fibers that can stretch and recoil through millions of cycles over a lifetime.
Elastin's amino acid composition is dominated by glycine, alanine, valine, and proline. Threonine is present at approximately 1.5-2% of residues. While this is lower than threonine's content in collagen, it is still meaningful because elastin synthesis occurs in a relatively narrow developmental window — most elastin in the human body is synthesized in utero and during the first decade of life, after which elastin turnover is extremely slow (estimated half-life of decades). The threonine that goes into elastin during childhood essentially has to last a lifetime.
This developmental window has clinical implications. Inadequate protein nutrition during pregnancy and early childhood — common in low-income populations globally — can compromise elastin synthesis, leading to reduced arterial elasticity, increased pulse-wave velocity, and elevated long-term cardiovascular risk. Population studies have correlated childhood undernutrition with adult arterial stiffness, and animal studies have shown that pregnant rats fed protein-restricted diets produce offspring with measurably reduced aortic elastin content.
Adults with established cardiovascular disease cannot rebuild elastin meaningfully — the synthetic machinery shuts down by the third decade. The clinical strategy is preventive: ensure adequate protein nutrition through pregnancy and childhood to maximize the elastin reserve laid down during the synthetic window. For more on cardiovascular tissue maintenance, see our Cardiology page and the Atherosclerosis deep-dive.
Tooth Enamel — Amelogenin and Enamelin
Tooth enamel is the hardest tissue in the human body, harder than bone, and is the most highly mineralized substance produced by any vertebrate (approximately 96% mineral by weight). Enamel is laid down by specialized cells called ameloblasts during tooth development — once the tooth has erupted, ameloblasts are lost and enamel cannot regenerate.
The mineral phase of enamel is hydroxyapatite, the same calcium phosphate mineral that forms the mineral phase of bone. The organic matrix that templates and guides hydroxyapatite crystal growth during enamel formation is composed of two principal proteins: amelogenin (approximately 90% of the developing enamel matrix protein content) and enamelin (about 5%), with ameloblastin and tuftelin filling smaller niches.
Amelogenin has a remarkable amino acid composition. Threonine is one of the most abundant residues, alongside proline, glutamine, leucine, and histidine. In some amelogenin isoforms, threonine reaches 10-15% of residues — making amelogenin one of the most threonine-rich proteins in the human body. The PYTYG motif (proline-tyrosine-threonine-tyrosine-glycine) appears repeatedly in the central region of amelogenin and is essential for the protein's ability to self-assemble into nanospheres that guide hydroxyapatite crystal growth.
The clinical implication is that adequate threonine nutrition during the years of tooth development (fetal life through age 13 or so, when the second molars complete enamel formation) is essential for proper enamel formation. Severe protein malnutrition during this window can produce enamel hypoplasia — thin, pitted, weak enamel that is more susceptible to caries and physical damage for the entire lifetime. This is part of why protein-energy malnutrition in childhood correlates with poor adult dental health long after nutritional rehabilitation.
Adults who have completed tooth development can no longer alter their enamel through nutrition. However, threonine remains important for:
- Saliva mucin glycoproteins — the protective film on tooth surfaces (the salivary pellicle) is built from threonine-rich mucin glycoproteins that buffer acid attacks and support remineralization
- Dentin and cementum maintenance — the mineralized tissues beneath enamel contain collagen as their primary organic matrix, and collagen turnover continues throughout life
- Periodontal connective tissue — the periodontal ligament that anchors teeth in the jaw is collagen-rich and turns over actively
Bone Matrix Proteins
Bone is approximately 65% mineral (hydroxyapatite) and 35% organic matrix. The organic matrix is dominated by type I collagen, which provides the tensile strength and flexibility that prevents bones from being brittle ceramics. Threonine's contribution to bone via type I collagen is therefore the same as in skin and tendon — approximately 2-3% of total residues.
Beyond collagen, several non-collagenous bone matrix proteins also contain meaningful threonine content:
- Osteocalcin — a small (49 amino acid) vitamin K-dependent protein produced by osteoblasts. Osteocalcin contains 2-3 threonine residues out of 49 (~4-6%), and is one of the most abundant non-collagenous proteins in bone. Threonine residues participate in the calcium-binding region that anchors osteocalcin to the hydroxyapatite mineral surface.
- Osteopontin — a larger phosphorylated glycoprotein that regulates bone remodeling. Osteopontin is approximately 8-10% threonine and contains multiple O-glycosylation sites at threonine residues that affect its binding to integrins on osteoclasts and osteoblasts.
- Bone sialoprotein — similar to osteopontin, contains threonine residues that are extensively glycosylated
The relevance to clinical practice: nutritional support for bone health emphasizes calcium, vitamin D, vitamin K2, magnesium, and protein. Within the protein category, ensuring adequate intake of complete proteins that provide collagen-building amino acids (glycine, proline, threonine, lysine) supports both the collagen scaffold and the non-collagenous matrix proteins. For more on bone health, see our pages on Vitamin D3, Vitamin K2, Calcium, and Osteoporosis.
Cartilage and Joint Proteoglycans
Articular cartilage is composed of type II collagen (about 90% of the collagen content), surrounded by a hydrated gel of proteoglycans — large protein-glycosaminoglycan complexes dominated by aggrecan. Aggrecan's protein core is decorated with hundreds of chondroitin sulfate and keratan sulfate side chains attached primarily to serine residues, with secondary attachment to threonine. The threonine-linked glycosaminoglycans contribute to the negative-charge density that draws water into cartilage and gives it its load-bearing properties.
Cartilage matrix turnover is slow under normal conditions but accelerates dramatically in osteoarthritis. The damaged cartilage in OA shows reduced proteoglycan content and reduced aggrecan glycosaminoglycan chain length, partly because the chondrocytes cannot synthesize new matrix fast enough to replace what is being degraded by matrix metalloproteinases. Adequate protein nutrition is one of many supportive measures in OA management, alongside weight management, low-impact exercise, and the role of collagen supplementation.
Glucosamine and chondroitin supplements work in part by providing the sugar substrates for glycosaminoglycan synthesis, but the protein backbone of aggrecan and the surrounding type II collagen still need to be synthesized from amino acids, and threonine is one of those required residues. For osteoarthritis management, see our Osteoarthritis page.
Wound Healing and Connective Tissue Repair
Wound healing proceeds through four overlapping phases: hemostasis, inflammation, proliferation, and remodeling. The proliferative phase, which begins around day 3 and peaks around day 7, is when fibroblasts produce massive quantities of new collagen and other extracellular matrix proteins to rebuild damaged tissue. This is the phase most sensitive to amino acid availability.
Wound collagen synthesis can transiently increase total collagen synthesis at the wound site to several times baseline. The amino acid demand is correspondingly high — protein-energy malnutrition delays wound healing measurably, with delayed wound closure, weaker scar tensile strength, and increased dehiscence risk in surgical wounds.
Within the protein nutrition question for wound healing, individual amino acids that have received specific attention include:
- Arginine — substrate for nitric oxide synthesis, also a substrate for proline (and thus hydroxyproline) production. Wound-healing nutritional formulas often emphasize arginine.
- Glutamine — fuel for rapidly dividing fibroblasts and immune cells
- Glycine and proline — direct substrates for collagen synthesis
- Threonine — required for both collagen synthesis and immunoglobulin production that protects against wound infection
- Lysine — substrate for collagen crosslinking via lysyl oxidase
Specialty wound-healing nutritional supplements often combine these amino acids with vitamin C (cofactor for prolyl and lysyl hydroxylases), zinc (cofactor for matrix metalloproteinases and lysyl oxidase), and vitamin A (epithelial differentiation). The evidence base for individual amino acids in isolation is weaker than the evidence for optimizing overall protein intake to 1.2-1.5 g/kg/day from high-biological-value sources during wound healing.
Crosslink Chemistry and Stability
Mature collagen fibrils gain their tensile strength from inter-molecular covalent crosslinks that bind adjacent collagen molecules into long arrays. The enzyme lysyl oxidase oxidizes specific lysine and hydroxylysine residues in the telopeptide regions of collagen to aldehydes, which then condense spontaneously with adjacent lysine, hydroxylysine, or histidine residues to form crosslinks.
While threonine itself is not directly oxidized into a crosslink, threonine residues in the surrounding sequence affect the local geometry, hydrogen bonding, and accessibility of the lysine and hydroxylysine residues that are oxidized. Sequence context matters — threonine-flanked lysines in some collagen subtypes form different crosslink patterns than alanine-flanked lysines, and these differences contribute to the distinct mechanical properties of skin collagen versus bone collagen versus tendon collagen.
The cofactors for crosslink formation are equally critical:
- Copper — lysyl oxidase is a copper-dependent enzyme. Copper deficiency produces brittle collagen, fragile blood vessels, and the classic Menkes disease phenotype in genetic copper transport defects.
- Vitamin C — cofactor for prolyl and lysyl hydroxylases. Vitamin C deficiency (scurvy) produces under-hydroxylated collagen with weak triple-helix stability.
- Pyridoxal phosphate (vitamin B6) — needed for some lysyl oxidase reactions
- Iron — cofactor for prolyl and lysyl hydroxylases (alongside vitamin C)
Adequate threonine in the substrate pool, plus adequate copper, vitamin C, iron, and vitamin B6 in the enzyme machinery, plus adequate glycine, proline, and lysine in the collagen sequence, all need to be present simultaneously for high-quality connective tissue formation. A deficiency in any one nutrient becomes the rate-limiting step. For more on these cofactors, see our pages on Copper, Vitamin C, and Vitamin B6.
Aging Connective Tissue
Connective tissue ages on a distinctive timeline. Elastin synthesis essentially ceases by the third decade of life, so the elastin that supports adult vasculature and skin elasticity is largely the elastin laid down by age 30. Collagen continues to be synthesized throughout life, but the synthesis rate slows by approximately 30% per decade after age 25, while the rate of accumulation of advanced glycation end products (AGEs) and other crosslink modifications accelerates.
The clinical signs of aging connective tissue include:
- Skin — thinning, loss of elasticity, increased wrinkling, slower wound healing, increased bruising from fragile dermal capillaries
- Cardiovascular — arterial stiffening (reduced elastin elasticity), elevated pulse-wave velocity, isolated systolic hypertension
- Skeletal — reduced bone matrix protein content alongside reduced mineral content, contributing to osteoporotic fracture risk
- Joint — cartilage thinning, reduced proteoglycan content, osteoarthritis
- Tendons and ligaments — reduced collagen turnover, increased AGE crosslinks producing stiffer but more brittle tissue, increased rupture risk in athletes over 40
While we cannot reverse the structural changes of aging connective tissue dramatically through nutrition alone, ensuring adequate amino acid supply (including threonine) for the ongoing — albeit reduced — collagen synthesis that continues throughout life is the foundation of supportive nutritional care. Population-level data suggests that older adults who maintain protein intake of 1.0-1.2 g/kg/day have less sarcopenia, less skin atrophy, and better wound healing than those who let protein intake drop into the 0.6-0.8 g/kg/day range that is unfortunately common in elderly populations.
Clinical Supplementation
Threonine as a standalone supplement for connective tissue support is uncommon. The much more common clinical approach is:
- Optimize total protein intake to 1.0-1.2 g/kg/day for sedentary adults, 1.2-1.6 g/kg/day for active adults, 1.2-2.0 g/kg/day for athletes and during recovery from injury or surgery. High-biological-value protein sources (eggs, fish, poultry, dairy, soy, quinoa) inherently provide the full essential amino acid profile including ample threonine.
- Hydrolyzed collagen / gelatin supplementation at 10-20 g/day provides large quantities of the collagen-building amino acids (glycine, proline, hydroxyproline) and meaningful threonine. Several clinical trials have shown improvement in joint pain, skin elasticity, and bone density with collagen peptide supplementation, though the strength of evidence varies by outcome.
- Bone broth as a whole-food source of collagen-building amino acids, traditionally consumed in many cultures for joint and gut health
- Vitamin and mineral cofactors — vitamin C (500-1,000 mg/day), zinc (15-30 mg/day), copper (2-3 mg/day) to ensure the enzyme machinery is well-fed
- Standalone L-threonine 100-500 mg/day only in specific clinical contexts (gut barrier repair, parenteral nutrition supplementation, occasional use during intense wound healing protocols when overall protein intake cannot be optimized)
For the related collagen-supporting nutrients, see our pages on Collagen, Bone Broth, Vitamin C, and Copper.
Cautions
- Whole-food protein remains the primary approach. Single amino acid supplements in isolation cannot substitute for adequate dietary protein. Threonine in particular is well-supplied by ordinary mixed diets that include eggs, dairy, fish, poultry, legumes, or soy.
- Renal impairment — isolated amino acid supplementation raises nitrogen load. Patients with advanced CKD should not self-supplement individual amino acids.
- Dental fluorosis and over-supplementation — childhood dental development requires balanced nutrition, not maximalism. Excessive supplementation of any single nutrient (fluoride, vitamin A, or amino acids) during the enamel-forming window can produce its own pathology.
- Connective tissue diseases — genetic connective tissue disorders (Ehlers-Danlos syndrome, Marfan syndrome, osteogenesis imperfecta) cannot be corrected by nutrition because the underlying defect is in the protein sequence, not in nutrient supply. Nutritional support is still important for general health, but unrealistic expectations of nutritional "cures" should be tempered.
- Aging — no amount of nutritional optimization will restore the elastin reserves that were laid down decades ago. Cardiovascular tissue maintenance becomes the rate-limiting step, not synthesis of new connective tissue.
Key Research Papers
- Shoulders MD, Raines RT (2009). Collagen structure and stability. Annual Review of Biochemistry. — PubMed
- Ramshaw JA, Shah NK, Brodsky B (1998). Gly-X-Y tripeptide frequencies in collagen: a context for host-guest triple-helical peptides. Journal of Structural Biology. — PubMed
- Eyre DR, Wu JJ (2005). Collagen cross-links. Topics in Current Chemistry. — PubMed
- Margolis HC et al. (2006). Role of macromolecular assembly of enamel matrix proteins in enamel formation. Journal of Dental Research. — PubMed
- Termine JD et al. (1980). Properties of dissociatively extracted fetal tooth matrix proteins. Journal of Biological Chemistry. (Foundational compositional analysis of amelogenin.) — PubMed
- Mithieux SM, Weiss AS (2005). Elastin. Advances in Protein Chemistry. — PubMed
- Avery NC, Bailey AJ (2006). The effects of the Maillard reaction on the physical properties and cell interactions of collagen. Pathologie Biologie. — PubMed
- Clark RA, Lin F, Greiling D, An J, Couchman JR (2004). Fibroblast invasive migration into fibronectin/fibrin gels requires a previously uncharacterized dermatan sulfate-CD44 proteoglycan. Journal of Investigative Dermatology. — PubMed
- Demling RH (2009). Nutrition, anabolism, and the wound healing process: an overview. Eplasty. — PubMed
- Zague V et al. (2011). Collagen hydrolysate intake increases skin collagen expression and suppresses matrix metalloproteinase 2 activity. Journal of Medicinal Food. — PubMed
- Clark KL et al. (2008). 24-Week study on the use of collagen hydrolysate as a dietary supplement in athletes with activity-related joint pain. Current Medical Research and Opinion. — PubMed
- Khatri M et al. (2021). The effects of collagen peptide supplementation on body composition, collagen synthesis, and recovery from joint injury and exercise: a systematic review. Amino Acids. — PubMed
PubMed Topic Searches
- PubMed: Threonine and collagen synthesis
- PubMed: Amelogenin composition
- PubMed: Elastin synthesis and nutrition
- PubMed: Collagen crosslinking
- PubMed: Collagen peptide supplementation
Connections
- Threonine Overview
- Threonine Benefits Hub
- Threonine for Gut Mucin
- Threonine for Immune Function
- Threonine for Liver Health
- Glycine (Collagen Partner)
- Proline (Collagen Partner)
- Lysine (Crosslinking)
- Collagen
- Bone Broth
- Vitamin C (Hydroxylase Cofactor)
- Copper (Lysyl Oxidase)
- Calcium
- Vitamin D3
- Vitamin K2
- Osteoporosis
- Osteoarthritis
- All Amino Acids