Threonine for Gut Mucin and the Intestinal Barrier
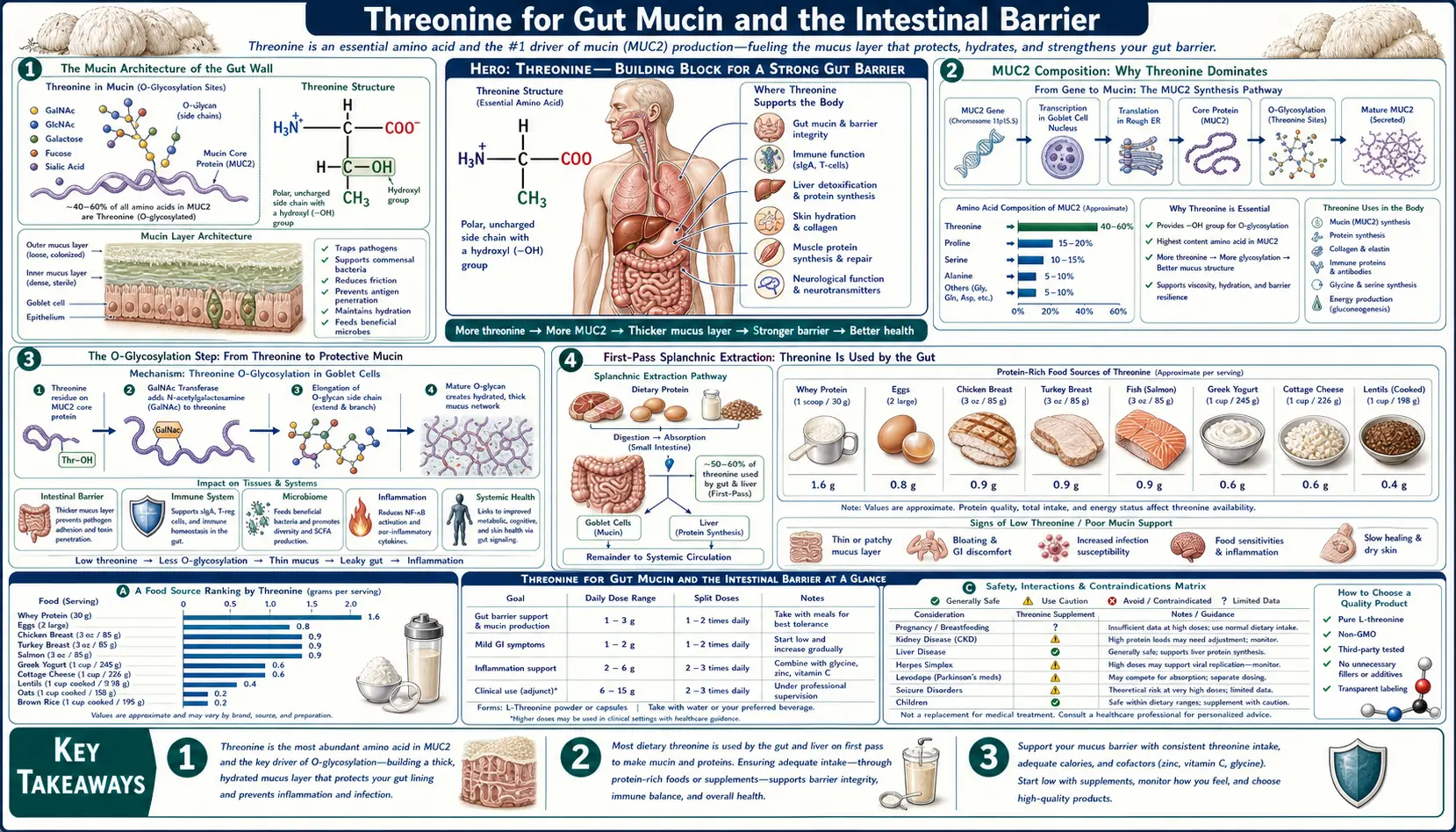
Threonine sits at the absolute foundation of intestinal-barrier biology. The dominant mucin secreted into the gut lumen — MUC2 — is roughly 30% threonine by amino-acid composition (with serine and proline filling the next two slots), making threonine the single most-required dietary amino acid for mucin synthesis. Without an intact MUC2 layer, the colon becomes directly exposed to its own bacterial inhabitants, which is the proximate mechanism that drives ulcerative colitis in MUC2-knockout mice and a recognized contributor to human inflammatory bowel disease. Wang and colleagues' classic 2010 piglet experiments showed that even a modest threonine restriction collapses goblet cell function, thins the mucus layer, and translates within days into bacterial translocation and weight loss. This page walks through the chemistry of mucin O-glycosylation, why threonine demand exceeds supply at the gut wall, and what the animal data tells human patients with IBD, ulcerative colitis, and post-infectious gut barrier dysfunction.
Table of Contents
- The Mucin Architecture of the Gut Wall
- MUC2 Composition: Why Threonine Dominates
- The O-Glycosylation Step
- First-Pass Splanchnic Extraction
- The Wang 2010 Piglet Studies
- The Two-Layer Colonic Mucus System
- MUC2 Knockout Mice and Spontaneous Colitis
- Translation to Human IBD and Ulcerative Colitis
- When Gut Bacteria Eat the Mucus Layer
- Clinical Implications and Practical Dosing
- Cautions
- Key Research Papers
- Connections
- Featured Videos
The Mucin Architecture of the Gut Wall
The lining of the human gastrointestinal tract is one of the largest interfaces between the body and the outside world — roughly 32 square meters of surface area when the villi and microvilli are accounted for, larger than a tennis court. This vast surface is covered continuously by a layer of mucus secreted by specialized goblet cells embedded in the epithelium. The mucus is not a passive coating. It is a structured gel built from large, heavily glycosylated proteins called mucins, and its integrity is the single most important determinant of whether the trillions of bacteria in the colonic lumen remain peacefully separated from the host or instead trigger destructive inflammation.
In the small intestine, the mucus is thinner and more permeable, allowing nutrient absorption while still providing a chemical and physical barrier to pathogens. In the colon, where the bacterial load is highest (1011-1012 organisms per gram of luminal content), the mucus is organized into two distinct layers: an inner, dense, stratified layer firmly adherent to the epithelium that is essentially bacteria-free, and an outer, looser layer that is colonized by commensal bacteria. The inner layer is renewed every hour by continuous secretion from goblet cells, and the outer layer is gradually solubilized by bacterial enzymes and shed into the stool.
The molecular building block of this mucus barrier is MUC2 (Mucin 2), a giant gel-forming mucin protein produced by goblet cells throughout the small and large intestine. Each MUC2 monomer is approximately 5,200 amino acids long, polymerizes head-to-head and tail-to-tail into massive multimers via cysteine disulfide bonds, and is so heavily glycosylated that 70-80% of its final mass is sugar rather than protein. This dense glycan coating creates the gel-like consistency that allows the mucus to trap pathogens and maintain its structural integrity under the mechanical stress of peristalsis.
MUC2 Composition: Why Threonine Dominates
The central region of MUC2 contains two large PTS (Proline-Threonine-Serine) tandem repeat domains. These regions are extraordinarily enriched in threonine, serine, and proline, and the hydroxyl groups of threonine and serine serve as the attachment points for the O-linked sugar chains that constitute the bulk of the molecule's mass.
Quantitative amino acid analysis of purified intestinal mucin yields a striking distribution:
- Threonine: approximately 28-32% of total amino acid residues
- Serine: approximately 16-20%
- Proline: approximately 13-18%
- All 17 other amino acids combined: the remaining 30-43%
For comparison, in the average tissue protein of the human body, threonine constitutes only about 5-6% of amino acid residues. The threonine content of MUC2 is therefore approximately five to six times higher than the body average. When the gut is producing mucin actively, the amino acid demand at the goblet cell is fundamentally skewed — the cell needs threonine at a rate that far exceeds what would be predicted from generic protein synthesis.
This explains an observation that has puzzled clinical nutritionists since the 1990s: dietary threonine seems to disappear into the gut. Stable-isotope tracer studies in young pigs (van Goudoever et al., Bertolo and Burrin) and in human infants and adults consistently show that 50-70% of dietary threonine is consumed by the splanchnic bed (gut plus liver) on first pass and never reaches systemic circulation. The disappearing threonine is largely going into mucin synthesis at the gut wall.
The O-Glycosylation Step
The reason threonine matters so much to mucin is the chemistry of O-glycosylation. After the MUC2 protein backbone is synthesized in the rough endoplasmic reticulum of the goblet cell, it moves to the Golgi apparatus, where a family of enzymes called polypeptide N-acetylgalactosaminyltransferases (GalNAc-Ts) transfer a single N-acetylgalactosamine sugar onto the hydroxyl oxygen of selected threonine and serine residues. This initial GalNAc attachment is the "Tn antigen" — the founding sugar of every O-linked glycan on the mucin.
From the Tn antigen, additional sugar units are added in a stepwise fashion by glycosyltransferases: galactose, N-acetylglucosamine, fucose, and terminal sialic acid or sulfate. The mature O-glycan chains extend outward from the protein backbone like the bristles of a bottle brush, creating the characteristic "Christmas tree" appearance of fully glycosylated mucin under high-resolution microscopy.
Crucially, only threonine and serine residues can serve as O-glycosylation sites — the chemistry requires a hydroxyl group on the amino acid side chain, and no other naturally occurring amino acid offers one in the right configuration (tyrosine has a hydroxyl but is positioned differently and is not used in mucin-type O-glycosylation). Because the PTS repeats of MUC2 are densely packed with threonine, almost every glycosylation site in the central region is a threonine. The polypeptide GalNAc-T family even shows substrate preference for threonine over serine in many sequence contexts (Wandall et al.), reinforcing the dominance of threonine residues as the chemical anchors for the entire glycan coat.
Without adequate threonine supply during MUC2 synthesis, two things go wrong. First, MUC2 backbones may be made with under-glycosylated PTS regions because some threonine sites are missing or substituted — the resulting mucin is structurally inferior and forms a thinner, less protective gel. Second, the total quantity of MUC2 produced drops because the goblet cell hits a threonine-limited synthesis ceiling. Either failure mode leaves the underlying epithelium exposed.
First-Pass Splanchnic Extraction
One of the most counterintuitive facts about threonine metabolism is the extraordinary fraction of dietary threonine that the gut and liver extract before it ever reaches the systemic circulation. Bertolo and colleagues at Memorial University documented this in piglet studies: when threonine is fed enterally (through the gut) versus parenterally (intravenously, bypassing the gut), enterally fed piglets need approximately 90% more threonine to maintain the same plasma concentration. The gap is the threonine being intercepted and used by the gut tissue itself, primarily for mucin synthesis.
In human studies, splanchnic extraction of dietary threonine has been estimated at 50-70% on first pass, the highest of any essential amino acid. By comparison, splanchnic extraction of lysine is about 30%, methionine 20%, and the branched-chain amino acids 20-25%. Threonine's extraction is uniquely large and is dominated by gut, not liver, consumption.
This has two clinical implications. First, recommended threonine intake values calibrated to systemic requirements (such as the WHO 15 mg/kg adult value) may underestimate true requirements in patients with increased gut mucin turnover — including IBD, post-infectious diarrhea, post-antibiotic gut recovery, and chronic NSAID-induced gut injury. Second, parenteral nutrition (TPN) formulations should provide more threonine than would be predicted by serum amino acid measurements, because TPN bypasses the splanchnic bed entirely and the gut still needs threonine to maintain its mucus barrier during the period of disuse atrophy.
The Wang 2010 Piglet Studies
The most-cited demonstration of threonine's role in maintaining the gut barrier comes from a series of experiments in young piglets by Wang and colleagues at China Agricultural University and parallel groups in Canada and France. Piglets are an unusually good model for human infant gastrointestinal physiology because of similarities in gut anatomy, mucin composition, and microbial colonization patterns.
Wang's 2010 study in the Journal of Nutrition fed weanling piglets diets containing 30%, 60%, or 100% of the standard threonine requirement for 14 days, then quantified gut mucin secretion, goblet cell density, and intestinal barrier function. Key findings:
- Piglets on 30% threonine showed a 40% reduction in jejunal MUC2 protein expression compared to the 100% group
- Goblet cell density in the small intestine fell by approximately 25% on the low-threonine diet
- The thickness of the firmly adherent mucus layer dropped by approximately 50%
- Intestinal permeability (measured as serum endotoxin and FITC-dextran flux) increased significantly
- Plasma threonine concentrations fell, but the gut mucin defect appeared before plasma threonine reached the conventional deficiency threshold — suggesting the gut is the first organ to suffer when threonine is marginally inadequate
Parallel work by Faure and colleagues at INRA in France used elegant 15N-threonine tracer infusion to directly quantify mucin synthesis rates under varying threonine intakes. They confirmed that gut mucin protein synthesis is exquisitely sensitive to dietary threonine supply — mucin synthesis rate falls within hours of inadequate intake, before any other tissue protein synthesis is affected.
The piglet data is mechanistic and dose-dependent. It is reasonable to extrapolate the principle (not the specific dose) to humans, particularly to the subgroups most likely to have inadequate threonine relative to gut demand: low-protein-intake patients, vegan diets that fail to combine complementary proteins adequately, IBD patients with increased mucin turnover, and parenterally fed patients on standard TPN formulations.
The Two-Layer Colonic Mucus System
The Gunnar Hansson laboratory at the University of Gothenburg, working through the 2000s and 2010s, established the modern understanding of the colonic mucus as a two-layer system. Using live-tissue imaging of mouse colon explants and bacterial visualization with FISH (fluorescent in situ hybridization), the Hansson group showed that:
- The inner mucus layer — approximately 50 micrometers thick in the mouse, perhaps 200 micrometers in the human — is firmly adherent to the epithelium and is essentially bacteria-free. It is built from densely packed, sulfated, and sialylated MUC2 polymers that form a near-impermeable physical barrier.
- The outer mucus layer — about 100 micrometers in mouse, much thicker in human — is structurally identical to the inner layer (same MUC2 backbone, same glycan coat) but has been partially proteolytically cleaved by bacterial enzymes. This loosens the gel, allowing commensal bacteria to colonize it. The outer layer functions as a colonization niche that confines bacteria to a defined region while the inner layer protects the epithelium.
Hansson's critical insight is that the inner layer is renewed continuously by goblet cell secretion at the base, while bacterial proteolysis erodes the outer surface. The system is in dynamic balance — if mucin secretion drops or proteolysis accelerates, the inner layer thins. When it thins to the point that bacteria can reach the epithelium, the host responds with inflammation, which is the precipitating event of ulcerative colitis.
Threonine availability is the rate-limiting nutritional input for the renewal side of this balance. When threonine is inadequate, MUC2 secretion drops, the inner layer cannot keep pace with bacterial erosion, and the colon transitions from a tolerated commensal state to an inflamed state.
MUC2 Knockout Mice and Spontaneous Colitis
Genetic ablation of MUC2 in mice produces one of the cleanest models of inflammatory bowel disease in modern immunology. Velcich et al. (2002, Science) created Muc2−/− mice by targeted gene disruption. The phenotype was dramatic:
- Muc2-null mice are viable and breed normally, but develop spontaneous colitis with bloody diarrhea, weight loss, and rectal prolapse by 6-8 weeks of age
- Histology shows dense bacterial colonization in direct contact with the colonic epithelium — no protective mucus layer exists
- Crypt architecture is disrupted, with goblet cells reduced and Paneth-like metaplasia appearing
- By 6-12 months, approximately 60% of Muc2-null mice develop colorectal adenomas and invasive cancer, demonstrating that chronic loss of mucus barrier drives both inflammation and oncogenesis
This experiment establishes that MUC2 is not merely beneficial but essential for colonic homeostasis — loss of MUC2 is sufficient to produce inflammatory bowel disease and ultimately colon cancer in an otherwise genetically normal mammal. The clinical implication is that anything which reduces MUC2 production (genetic, nutritional, pharmacological) should be expected to push the colon toward inflammation. Threonine deficiency is the most common and easily corrected nutritional driver of MUC2 production failure.
Subsequent work has shown that Muc2 expression is also reduced in human ulcerative colitis biopsies (van der Sluis et al.), and rare germline mutations in MUC2 itself or in the enzymes responsible for mucin O-glycosylation are associated with familial inflammatory bowel disease phenotypes. The MUC2 mucus barrier is, in retrospect, one of the central nodes in IBD pathogenesis.
Translation to Human IBD and Ulcerative Colitis
Several lines of evidence link impaired mucin biology in humans to inflammatory bowel disease, with implications for threonine nutrition as part of the broader treatment landscape.
Reduced mucus thickness in active ulcerative colitis. Pullan and colleagues (1994, Gut) measured colonic mucus thickness in patients with active UC, patients in remission, and healthy controls using a sensitive ex vivo technique. The thickness in active disease was reduced by approximately 50% compared to controls. Patients in remission had intermediate values. This finding has been replicated multiple times.
Reduced MUC2 expression and altered glycosylation. Biopsy studies in UC patients show reduced MUC2 protein expression, often with abnormal glycan structures (truncated O-glycans, reduced sulfation). These changes correlate with disease activity and inversely with the depth of clinical remission.
Plasma threonine in active IBD. Several studies have documented reduced plasma threonine concentrations in patients with active Crohn's disease and ulcerative colitis. The mechanism is multifactorial — reduced intake due to anorexia, increased splanchnic extraction to compensate for accelerated mucin turnover, malabsorption, and increased catabolic losses in inflammation. The net result is that IBD patients are at meaningfully increased risk for threonine inadequacy relative to their elevated demand.
Threonine supplementation in animal models of colitis. Multiple groups have shown that dietary threonine supplementation reduces severity of chemically induced colitis (DSS, TNBS) in rats and mice, partly by restoring mucin synthesis and partly through reduction of pro-inflammatory cytokine expression. Whether this translates to human IBD remains under investigation — there are no large randomized trials of isolated threonine supplementation in UC or Crohn's. Practical nutritional management in IBD focuses on optimizing total protein intake from high-biological-value sources, which inherently provides adequate threonine.
For a broader treatment of inflammatory bowel disease pathogenesis and management, see our Inflammatory Bowel Disease page and the Crohn's Disease and Ulcerative Colitis deep-dives.
When Gut Bacteria Eat the Mucus Layer
A particularly elegant aspect of mucin biology that has emerged in the last fifteen years is that certain gut bacteria specialize in degrading the mucus layer as their primary carbon source. Akkermansia muciniphila, a Verrucomicrobia species first cultured by Derrien and colleagues in 2004, is the best-studied mucin-degrading bacterium and lives almost exclusively at the outer surface of the colonic mucus layer.
This sounds destructive, but it is actually a stable and beneficial relationship in the healthy gut. Akkermansia trims back the outer mucus layer at a rate that the goblet cells can match through new secretion, releasing oligosaccharides that other beneficial commensals (including butyrate-producing Firmicutes) consume. The result is a healthy steady-state turnover of the mucus layer. Akkermansia abundance correlates positively with metabolic health, reduced obesity, and reduced inflammation in dozens of cohort studies.
The problem arises when goblet cell secretion fails to match bacterial degradation. If MUC2 production drops — whether from threonine inadequacy, low-fiber diet that paradoxically increases mucin-degrading bacterial expansion, or genetic insufficiency — the inner mucus layer thins and bacteria gain access to the epithelium. Desai and colleagues (2016, Cell) showed in gnotobiotic mice fed a low-fiber diet that mucin-degrading bacteria preferentially shifted to consuming the host mucus layer, thinning it to the point of bacterial-epithelial contact and inflammation. Adding fiber back to the diet restored the mucus layer within days.
The take-home message is that the gut mucus barrier is the product of a three-way balance: dietary threonine (substrate for mucin synthesis), dietary fiber (alternative substrate that keeps mucin-degrading bacteria from eating too much of the host mucus), and adequate goblet cell health (mucin secretion capacity). Disruption of any one of these inputs — common in modern diets — predisposes to barrier failure and the chronic low-grade inflammation that follows.
Clinical Implications and Practical Dosing
For most healthy adults eating a varied diet with adequate protein, threonine intake comfortably exceeds the WHO recommendation of 15 mg/kg/day. The clinical situations where threonine supply may become limiting for mucin production are specific:
- Active IBD with elevated mucin turnover — threonine demand is increased and intake is often reduced. Optimize overall protein intake to 1.2-1.5 g/kg/day from high-quality sources (eggs, fish, poultry, dairy if tolerated, plus targeted plant proteins). Whey protein concentrate is particularly threonine-dense.
- Post-antibiotic gut recovery — the period after broad-spectrum antibiotics is when the mucin layer is thinnest and rebuilding it is a priority. Optimize protein, add fermented foods, and consider a short course of L-threonine 500-1,000 mg/day in divided doses for 4-8 weeks if intake is uncertain.
- Parenteral nutrition — standard TPN amino acid solutions provide threonine, but the gut is bypassed and atrophies during prolonged TPN. Adding small amounts of enteral nutrition ("trophic feeds") maintains gut mucin production better than threonine dosing alone.
- Strict vegan diets without protein combining — threonine intake from grain-only diets can be marginal because grains are limiting in threonine. Adding legumes, soy protein, quinoa, or seeds at every meal corrects this. Pure rice-based diets are particularly threonine-low and should be supplemented.
- Bariatric surgery patients — reduced food volume and altered absorption can drop threonine intake below requirements. Lifelong attention to protein intake (60-80 g/day minimum from high-biological-value sources) is standard.
Standalone L-threonine supplementation is not commonly necessary if total protein intake is adequate. When used, typical clinical doses are 100-500 mg three times daily between meals. Some clinicians use higher doses (1-2 g/day) for short-term gut healing protocols, divided into 3-4 doses. Doses above 2 g/day are rarely used and risk amino acid imbalance — isolated dosing of any single essential amino acid can interfere with absorption of others at the gut wall.
Cautions
- Threonine alone is not a treatment for IBD. Inflammatory bowel disease is multifactorial. Threonine optimization is supportive nutritional care, not a substitute for medical management with mesalamine, biologics, immunomodulators, or surgical care as appropriate.
- Renal impairment — isolated amino acid supplementation increases nitrogen load. Patients with advanced chronic kidney disease should not use high-dose individual amino acid supplements without nephrology guidance.
- Phenylketonuria and other inborn errors of amino acid metabolism — threonine catabolism feeds into the threonine-aldolase or threonine-dehydratase pathways, neither of which is affected by classical PKU. Threonine is generally safe in PKU patients but specialist guidance is warranted for any amino acid supplementation in inborn errors.
- Pregnancy — whole-food protein sources are preferred. There is no established role for isolated threonine supplementation in pregnancy.
- The mucus barrier needs more than threonine. Mucin O-glycosylation requires UDP-GalNAc, UDP-Gal, UDP-GlcNAc, GDP-fucose, and CMP-sialic acid, all of which depend on adequate carbohydrate and intermediate metabolism. Severe carbohydrate restriction (long-term strict ketogenic diets, prolonged fasting) reduces the substrates available for glycan synthesis even when threonine is plentiful.
Key Research Papers
- Wang X, Qiao S, Yin Y, Yue L, Wang Z, Wu G (2010). Threonine-deficient diet affects gut mucin synthesis and reduces intestinal barrier function in piglets. Journal of Nutrition. — PubMed
- Velcich A et al. (2002). Colorectal cancer in mice genetically deficient in the mucin Muc2. Science. — PubMed
- van der Sluis M et al. (2006). Muc2-deficient mice spontaneously develop colitis, indicating that MUC2 is critical for colonic protection. Gastroenterology. — PubMed
- Johansson MEV et al. (2008). The inner of the two Muc2 mucin-dependent mucus layers in colon is devoid of bacteria. PNAS. — PubMed
- Hansson GC (2012). Role of mucus layers in gut infection and inflammation. Current Opinion in Microbiology. — PubMed
- Pullan RD et al. (1994). Thickness of adherent mucus gel on colonic mucosa in humans and its relevance to colitis. Gut. — PubMed
- Bertolo RF et al. (1998). Threonine requirement of neonatal piglets receiving total parenteral nutrition is considerably lower than that of piglets receiving an identical diet intragastrically. Journal of Nutrition. — PubMed
- Faure M et al. (2006). Specific amino acids increase mucin synthesis and microbiota in dextran sulfate sodium-treated rats. Journal of Nutrition. — PubMed
- Derrien M et al. (2004). Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. International Journal of Systematic and Evolutionary Microbiology. — PubMed
- Desai MS et al. (2016). A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell. — PubMed
- van Goudoever JB et al. (2000). Adaptive regulation of intestinal lysine metabolism. PNAS. (Methodologic basis for splanchnic-extraction studies that later targeted threonine.) — PubMed
- Wandall HH et al. (1997). Substrate specificities of three members of the human UDP-N-acetyl-alpha-D-galactosamine:Polypeptide N-acetylgalactosaminyltransferase family. Journal of Biological Chemistry. — PubMed
- Bansil R, Turner BS (2018). The biology of mucus: composition, synthesis and organization. Advanced Drug Delivery Reviews. — PubMed
PubMed Topic Searches
- PubMed: Threonine and intestinal mucin
- PubMed: MUC2 and IBD
- PubMed: Goblet cells and mucus barrier
- PubMed: Threonine splanchnic extraction
- PubMed: Akkermansia and mucus
Connections
- Threonine Overview
- Threonine Benefits Hub
- Threonine for Collagen
- Threonine for Immune Function
- Threonine for Liver Health
- Glutamine (Enterocyte Fuel)
- Serine (Co-residue in PTS Repeats)
- Proline (Third PTS Residue)
- Glycine
- Inflammatory Bowel Disease
- Ulcerative Colitis
- Crohn's Disease
- Leaky Gut Syndrome
- SIBO
- Gut-Brain Axis
- Bone Broth
- Eggs
- All Amino Acids