Vitamin B4 (Adenine), DNA and RNA Synthesis
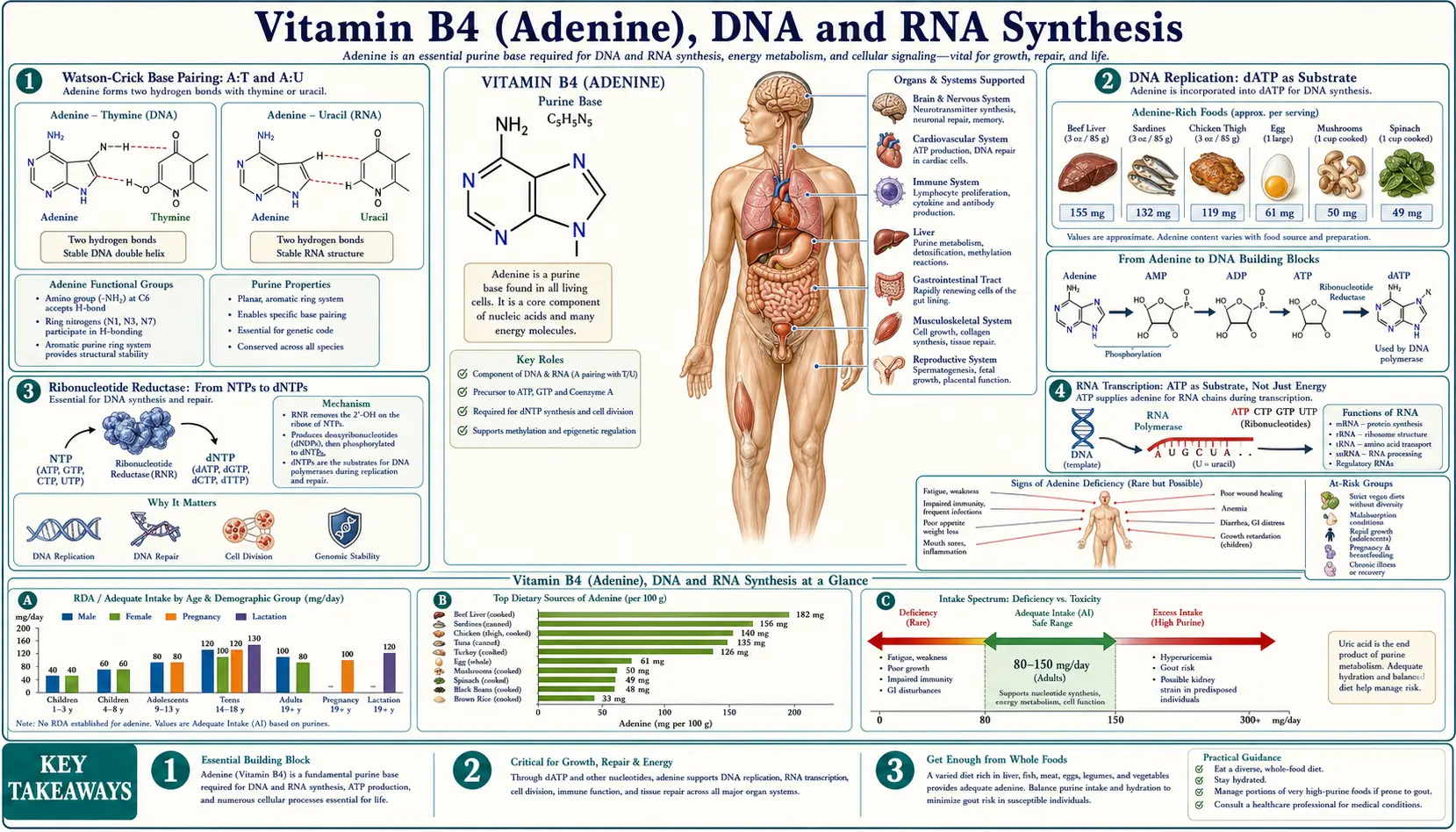
Adenine is one of the four nitrogenous bases that, together with thymine, guanine, and cytosine (DNA) or uracil instead of thymine (RNA), constitute the universal genetic alphabet. The Watson-Crick A:T base pair (two hydrogen bonds) and A:U pair (RNA) hold the nucleic acid double helix together and provide the templating geometry that makes accurate replication and transcription possible. Every cell division duplicates roughly 6 × 109 bases per diploid genome, of which approximately one quarter — 1.5 billion residues — are adenine. Every transcription event further consumes ATP as substrate (RNA polymerase uses ATP, GTP, CTP, and UTP as direct building blocks, not the deoxynucleotides used by DNA polymerase). This deep-dive walks through the enzymology, the connection to chemotherapy drug design, and the special role of adenine modifications in epigenetics.
Table of Contents
- Watson-Crick Base Pairing: A:T and A:U
- DNA Replication: dATP as Substrate
- Ribonucleotide Reductase: From NTPs to dNTPs
- RNA Transcription: ATP as Substrate, Not Just Energy
- Proliferating Tissues and Purine Demand
- Antifolate Chemotherapy: Methotrexate and Pemetrexed
- Purine Analog Chemotherapy: 6-MP, 6-TG, Fludarabine, Cladribine
- Antiviral Nucleoside Analogs: Acyclovir, Tenofovir, Remdesivir
- N6-Methyladenosine (m6A) and RNA Epigenetics
- Key Research Papers
- Connections
- Featured Videos
Watson-Crick Base Pairing: A:T and A:U
The 1953 Watson-Crick model of the DNA double helix proposed two principles that have proved essentially correct in every detail explored since: antiparallel strands (one running 5'-to-3', the other 3'-to-5') and specific base pairing across the helix (adenine pairs with thymine, guanine with cytosine). The pairing is enforced by hydrogen-bonding geometry — A:T pairs share two hydrogen bonds, G:C pairs share three — and by the constraint that pairing geometry must fit within a uniform helical diameter, which only purine-pyrimidine pairs achieve.
Adenine in the A:T pair acts as a hydrogen-bond donor at the 6-amino group (N6-H…O4=C of thymine) and as a hydrogen-bond acceptor at N1 (N1…HN3 of thymine). In RNA, adenine pairs with uracil through the same geometry — uracil is identical to thymine except for the absence of a methyl group at C5, and the methyl difference does not perturb the base-pairing chemistry.
The G:C pair is intrinsically more stable than A:T because it adds a third hydrogen bond (N4-H…O6, N1-H…N3, and N2-H…O2 of cytosine). This difference produces measurable consequences: A:T-rich DNA regions melt at lower temperatures than G:C-rich regions (the basis of PCR primer design and melting-curve analysis), and the AT-rich origins of bacterial DNA replication are deliberately easier to unwind than the surrounding genome.
Non-Watson-Crick pairings occur under specialized conditions. Hoogsteen base pairing (in which adenine flips to pair with thymine through its N7 face instead of N1) is found in triple-helical DNA structures, in certain transcription factor binding sites, and in some tRNA tertiary structures. Wobble base pairing (G:U) is essential for the genetic code, allowing a single tRNA to recognize multiple codons differing only in the third position.
DNA Replication: dATP as Substrate
DNA replication is catalyzed by DNA polymerases that copy a template strand by adding deoxyribonucleoside triphosphates (dNTPs) one at a time to the growing 3'-end of the daughter strand. The four dNTPs — dATP, dTTP, dGTP, dCTP — are the direct substrates. The polymerase reaction releases pyrophosphate, which is rapidly hydrolyzed by pyrophosphatase to two phosphate groups, making the overall reaction thermodynamically irreversible.
Eukaryotic DNA replication uses a coordinated set of polymerases: Pol alpha primes both leading and lagging strands with short RNA-DNA primers; Pol epsilon performs continuous leading-strand synthesis; Pol delta performs discontinuous lagging-strand (Okazaki fragment) synthesis. All four use the same dNTP pool, and all four require accurate fidelity through proofreading via 3'-to-5' exonuclease domains.
The fidelity of replication is roughly 10-7 errors per nucleotide before proofreading, improved to 10-9 with proofreading and roughly 10-10 after mismatch repair acts on the daughter strand. Adenine is the most commonly miscoded base — A:G and A:C mispairs are the most frequent polymerase errors — because adenine's hydrogen-bonding profile permits unusually tolerant non-Watson-Crick pairing geometries.
The dNTP pool sizes are tightly regulated. Cells maintain dATP at the lowest concentration of the four dNTPs (roughly 25 μM, vs. ~70 μM for dTTP). The reason is that elevated dATP feedback-inhibits ribonucleotide reductase, the enzyme that produces all four dNTPs. Pathologic dATP elevation — as occurs in ADA-SCID — collapses dNTP supply, blocks DNA synthesis, and kills proliferating lymphocytes preferentially.
Ribonucleotide Reductase: From NTPs to dNTPs
Ribonucleotide reductase (RNR) is the single enzyme responsible for converting all four ribonucleoside diphosphates (ADP, GDP, CDP, UDP) to the corresponding 2'-deoxyribonucleoside diphosphates (dADP, dGDP, dCDP, dUDP). It accomplishes the reduction by replacing the 2'-hydroxyl group of ribose with a hydrogen atom, using a remarkable free-radical mechanism that involves a stable tyrosyl radical (in the R2 subunit) coupled through a long-range electron transfer pathway to the active site (in the R1 subunit).
RNR is allosterically regulated to balance the dNTP pools. The activity site (binding ATP or dATP) controls overall flux: ATP turns the enzyme on, dATP turns it off. The substrate specificity site (binding dATP, dTTP, dGTP) determines which ribonucleotide diphosphate is preferentially reduced. The result is a coordinated regulation that prevents any single dNTP from accumulating to toxic levels while ensuring sufficient supply of each for accurate DNA synthesis.
RNR is the molecular target of several anticancer drugs. Hydroxyurea quenches the tyrosyl radical, shutting down dNTP production and blocking DNA synthesis — used in sickle-cell disease (where it also raises fetal hemoglobin), polycythemia vera, essential thrombocythemia, and chronic myeloid leukemia. Gemcitabine, a deoxycytidine analog, is phosphorylated to gemcitabine diphosphate which inhibits RNR; it is used in pancreatic, lung, and bladder cancers. Cladribine and fludarabine, purine analogs discussed below, also indirectly suppress RNR.
RNA Transcription: ATP as Substrate, Not Just Energy
RNA polymerases use ribonucleoside triphosphates (ATP, GTP, CTP, UTP) directly as substrates — not the deoxynucleotides used by DNA polymerase. In RNA synthesis, ATP serves both roles simultaneously: it provides the energy for the polymerization reaction (via pyrophosphate release and hydrolysis), and it provides the adenosine residue that is incorporated into the growing RNA strand.
The three eukaryotic RNA polymerases have distinct functions. RNA polymerase I transcribes the rRNA precursor in the nucleolus, producing ~80% of total cellular RNA by mass. RNA polymerase II transcribes mRNAs and most regulatory non-coding RNAs — the workhorse of gene expression. RNA polymerase III transcribes tRNAs, 5S rRNA, and a handful of small regulatory RNAs.
Each transcription event consumes ATP not only as substrate for adenosine incorporation but also for several auxiliary processes: capping (which adds a 7-methylguanosine to the 5' end via a triphosphate linkage), splicing (which uses ATP-dependent helicases to remodel the spliceosome), polyadenylation (which adds a long stretch of adenosine residues to the 3' end using ATP as direct substrate), and nuclear export (which uses ATP-driven motors).
The poly-A tail is one of the largest single ATP sinks per transcript: a typical mRNA carries 150–250 adenosine residues at its 3' end, added post-transcriptionally by poly-A polymerase. The poly-A tail protects the mRNA from 3'-exonuclease degradation, promotes nuclear export, and stimulates translation through interaction with poly-A binding protein. The eventual shortening of the poly-A tail (deadenylation) triggers decapping and 5'-to-3' exonuclease degradation, controlling mRNA half-life.
Proliferating Tissues and Purine Demand
Cells that divide rapidly have correspondingly high demand for dNTPs and NTPs, and therefore for purine synthesis capacity. The most proliferative tissues in the adult body are bone marrow (which produces ~2 × 1011 blood cells per day), gut crypts (which renew the entire intestinal epithelium every 4–5 days), and hair follicles. In children, growing tissues including bone, skin, and muscle are similarly purine-demanding. In pregnancy, embryonic tissue is the most purine-demanding tissue in the body, period.
The high purine demand of these tissues is also their vulnerability. Most cytotoxic chemotherapy drugs work by interfering with DNA or RNA synthesis, and their predictable side-effect profile — myelosuppression, mucositis, alopecia — reflects the collateral damage to the same proliferating tissues that drove their efficacy against the tumor. Pregnancy is an absolute contraindication for most chemotherapy in the first trimester because the embryonic tissue is even more vulnerable than the tumor.
The reverse principle is also exploited clinically. Lymphocytes (particularly activated, proliferating T and B cells) have unusually high de novo purine synthesis demand and limited salvage capacity. This is why purine-pathway-targeting drugs (mycophenolate mofetil, azathioprine) are useful as immunosuppressants in transplantation, IBD, and autoimmune disease — the lymphocyte selectivity comes from the tissue's specific dependence on the de novo pathway.
The selective vulnerability of fast-proliferating cells to purine starvation also explains why dietary purine restriction has clinical relevance in only a narrow set of conditions (gout, tumor lysis syndrome, certain inborn errors). For most patients, the body's adaptive upregulation of de novo synthesis fully compensates for reduced dietary intake.
Antifolate Chemotherapy: Methotrexate and Pemetrexed
Methotrexate is the prototype antifolate. It binds dihydrofolate reductase (DHFR) with affinity ~1000-fold greater than the natural substrate dihydrofolate, depleting the reduced-folate pool. Without tetrahydrofolate, the one-carbon transfer reactions required for de novo purine synthesis (formylation of glycinamide ribonucleotide and aminoimidazolecarboxamide ribonucleotide by formyl-THF) stall, and de novo pyrimidine synthesis stalls at thymidylate synthesis. Cells starved of dNTPs cannot replicate DNA and die by apoptosis if they cannot exit the cell cycle.
Methotrexate is used in: acute lymphoblastic leukemia (high-dose with leucovorin rescue), choriocarcinoma (where it was the first solid tumor cured by chemotherapy), rheumatoid arthritis (low weekly dose, anti-inflammatory mechanism still incompletely understood), psoriasis, Crohn disease, and ectopic pregnancy. Folate supplementation (1 mg/day folic acid) reduces methotrexate toxicity without compromising efficacy in low-dose rheumatologic use.
Pemetrexed is a newer antifolate that inhibits DHFR plus thymidylate synthase plus glycinamide ribonucleotide formyltransferase — a more selective hit to purine and thymidylate synthesis. Used in non-small-cell lung cancer and mesothelioma. The label requires concurrent vitamin B12 and folate supplementation to reduce hematologic toxicity.
Pralatrexate is used in peripheral T-cell lymphoma. Trimethoprim selectively inhibits bacterial DHFR (with much lower affinity for human DHFR), and is the antifolate component of the antibiotic combination trimethoprim-sulfamethoxazole.
Purine Analog Chemotherapy: 6-MP, 6-TG, Fludarabine, Cladribine
Purine analogs are structural mimics of adenine, guanine, or hypoxanthine that, once phosphorylated to the nucleotide form by salvage enzymes (HGPRT, deoxycytidine kinase) or by viral kinases, are incorporated into DNA or interfere with purine metabolism.
6-mercaptopurine (6-MP) and 6-thioguanine (6-TG) are hypoxanthine and guanine analogs developed by Gertrude Elion in the 1950s (Nobel Prize 1988). After salvage by HGPRT, the resulting thio-IMP and thio-GMP both feedback-inhibit GPAT (suppressing further de novo purine synthesis) and incorporate into DNA where they disrupt replication. Both are mainstays of pediatric acute lymphoblastic leukemia maintenance therapy.
Azathioprine is a 6-MP prodrug that releases 6-MP non-enzymatically in vivo. Used as an immunosuppressant in inflammatory bowel disease, lupus, vasculitis, transplantation. TPMT genotyping before initiation is now standard because patients with low or absent TPMT activity accumulate toxic thio-nucleotides and develop severe myelosuppression.
Fludarabine is a fluorinated adenosine analog that, after phosphorylation, inhibits DNA polymerase, ribonucleotide reductase, and DNA primase. The triphosphate also incorporates into DNA. Workhorse drug for chronic lymphocytic leukemia.
Cladribine (2-chlorodeoxyadenosine) is a deoxyadenosine analog that is resistant to deamination by adenosine deaminase. After phosphorylation by deoxycytidine kinase, it accumulates as cladribine triphosphate and incorporates into DNA. Highly effective for hairy cell leukemia (90% complete remission rate with a single 7-day course) and approved for multiple sclerosis.
Clofarabine is a second-generation purine analog used in relapsed pediatric ALL. Nelarabine is a guanine analog used in T-cell ALL and T-cell lymphoblastic lymphoma.
Antiviral Nucleoside Analogs: Acyclovir, Tenofovir, Remdesivir
Antiviral nucleoside analogs exploit the same selective-incorporation principle as purine-analog chemotherapy, but achieve selectivity for virus-infected cells through differential phosphorylation by viral enzymes.
Acyclovir is a guanosine analog that, in uninfected cells, is barely phosphorylated. In herpesvirus-infected cells, the viral thymidine kinase phosphorylates acyclovir efficiently to acyclovir monophosphate; cellular kinases then complete the phosphorylation to acyclovir triphosphate, which incorporates into viral DNA and terminates elongation (the missing 3'-OH on the acyclic sugar prevents further nucleotide addition). This selective phosphorylation gives acyclovir a roughly 1000-fold selectivity for infected cells.
Tenofovir is an acyclic adenosine analog used in HIV and hepatitis B. The phosphonate group bypasses the rate-limiting first phosphorylation step required by classical nucleoside analogs, allowing tenofovir to be activated even in cells lacking the appropriate kinase. The triphosphate inhibits HIV reverse transcriptase and HBV polymerase.
Remdesivir is a 1'-cyano adenosine analog originally developed for Ebola virus, repurposed during the 2020 pandemic for emergency use, and subsequently FDA-approved for hospitalized patients with severe respiratory viral infection. It targets the viral RNA-dependent RNA polymerase and acts as a delayed chain terminator.
Adefovir (chronic hepatitis B), didanosine (HIV, now rarely used due to toxicity), abacavir (HIV, requires HLA-B*57:01 screening to prevent hypersensitivity), and entecavir (chronic hepatitis B) are additional clinically used purine nucleoside analog antivirals.
N6-Methyladenosine (m6A) and RNA Epigenetics
The most abundant chemical modification of mRNA in mammalian cells is N6-methyladenosine (m6A) — the methylation of adenine's 6-amino group. Roughly 0.1–0.4% of all adenine residues in mRNA carry the m6A modification, totaling roughly three modifications per typical mRNA. The distribution is not random — m6A clusters near stop codons, in 3' untranslated regions, and in long internal exons.
The m6A modification is installed by a multi-subunit "writer" complex (METTL3, METTL14, WTAP), removed by "erasers" (FTO, ALKBH5), and recognized by "reader" proteins (YTHDF1-3, YTHDC1-2, IGF2BP1-3). This writer-eraser-reader architecture parallels the writer-eraser-reader paradigm of histone modifications and DNA methylation — an "RNA epigenetics" layer that regulates mRNA splicing, stability, translation efficiency, and nuclear export.
Mutations in m6A pathway components are associated with cancer, obesity, and neurological disease. FTO (one of the m6A erasers) was originally identified as the strongest common-variant obesity risk locus in humans, before its enzymatic function as an m6A demethylase was discovered. METTL3 is overexpressed in acute myeloid leukemia and is being targeted by small-molecule METTL3 inhibitors in clinical trials.
Other adenine RNA modifications include inosine (produced by adenosine deaminase acting on RNA, ADAR; converts A to I, which is read as G during translation — a form of post-transcriptional codon recoding), 2'-O-methyladenosine (a common modification in rRNA and snoRNA), and N1-methyladenosine (found in tRNA and a subset of mRNAs).
The DNA-side analog of m6A is N6-methyl-2'-deoxyadenosine (6mA), which has been detected at low levels in mammalian DNA and may serve regulatory roles — though the field is still controversial about whether mammalian 6mA reflects genuine endogenous DNA modification or contamination from RNA breakdown products.
Key Research Papers
- Watson JD and Crick FH, molecular structure of nucleic acids (1953 Nature) — PubMed: Watson-Crick 1953
- Kornberg A, enzymatic synthesis of DNA (1956 Nobel work) — PubMed: Kornberg DNA polymerase
- Nordlund P and Reichard P, ribonucleotide reductase mechanism — PubMed: RNR mechanism
- Roeder RG, three eukaryotic RNA polymerases (I, II, III) — PubMed: Roeder three polymerases
- Wahle E and Keller W, mRNA polyadenylation mechanism and poly-A binding protein — PubMed: Polyadenylation
- Elion GB and Hitchings GH, 6-mercaptopurine design and Nobel work — PubMed: Elion-Hitchings
- Schaeffer HJ and Gertner M, acyclovir mechanism and selectivity — PubMed: Acyclovir mechanism
- Holy A, tenofovir and acyclic nucleoside phosphonates — PubMed: Tenofovir history
- Beigel JH, remdesivir for COVID-19 (ACTT-1 trial NEJM 2020) — PubMed: ACTT-1 remdesivir
- Dominissini D, topology of m6A methylome in human and mouse transcriptomes — PubMed: m6A methylome
- Jia G, FTO as the first identified m6A demethylase — PubMed: FTO m6A demethylase
- Liu J, METTL3-METTL14 complex as m6A writer — PubMed: METTL3/14 writer
Connections
- Vitamin B4 Benefits (Hub)
- Vitamin B4 (Main Page)
- Purine Metabolism
- Cellular Energy and ATP
- Modern Status and Food Sources
- Vitamin B9 (Folate)
- Vitamin B12
- Vitamin B6
- Leukemia
- Lymphoma
- Sickle Cell Disease
- Rheumatoid Arthritis
- Immune Boosting
- CBC (Bone Marrow Function)
- All Vitamins