Vitamin B4 (Adenine), Cellular Energy and ATP
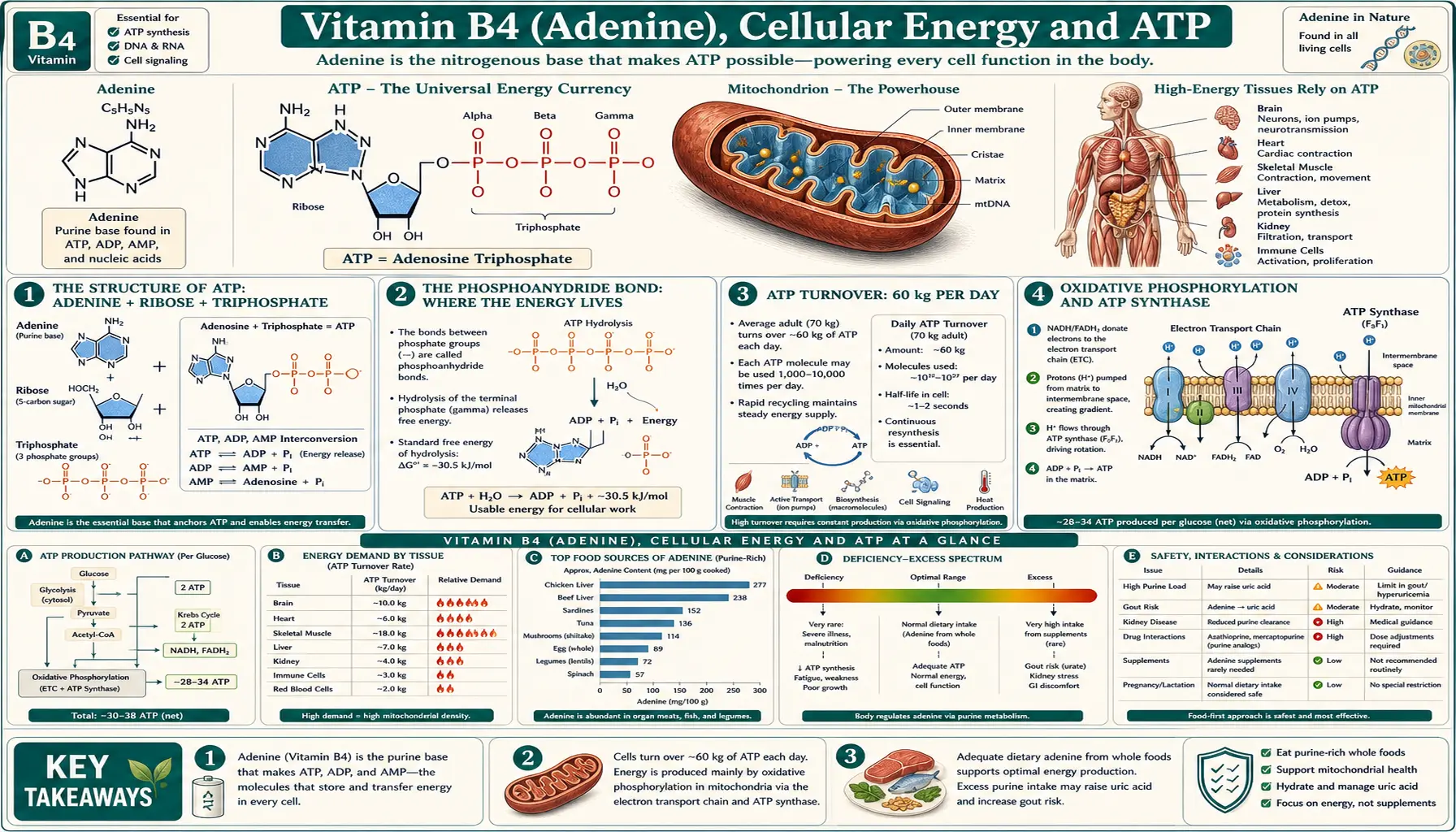
Adenosine triphosphate (ATP) is the universal energy currency of cellular metabolism, and its nitrogenous base — adenine, the molecule once called Vitamin B4 — is the recognition handle that ATP-using enzymes evolved to bind. A typical 70 kg human adult turns over approximately 60 kg of ATP per day, recycling the same pool of roughly 250 g of adenine nucleotides through ADP↔ATP cycles hundreds of times daily. The same adenine scaffold appears in nicotinamide adenine dinucleotide (NAD/NADH), nicotinamide adenine dinucleotide phosphate (NADP/NADPH), flavin adenine dinucleotide (FAD/FADH2), coenzyme A (acetyl-CoA, succinyl-CoA, malonyl-CoA), S-adenosylmethionine (SAM), and the cyclic second messengers cAMP and cGMP. Without adenine, almost no enzymatic reaction in central metabolism could proceed.
Table of Contents
- The Structure of ATP: Adenine + Ribose + Triphosphate
- The Phosphoanhydride Bond: Where the Energy Lives
- ATP Turnover: 60 kg per Day
- Oxidative Phosphorylation and ATP Synthase
- Substrate-Level Phosphorylation in Glycolysis and the TCA Cycle
- Adenine as Cofactor Scaffold: NAD, FAD, CoA, SAM
- Adenine in Cellular Signaling: cAMP, AMPK, and Adenosine Receptors
- Muscle Energetics: Creatine Phosphate Buffer
- Mitochondrial Disease and ATP Production Failure
- Key Research Papers
- Connections
- Featured Videos
The Structure of ATP: Adenine + Ribose + Triphosphate
Adenosine triphosphate consists of three covalently joined parts. The adenine base — a fused two-ring nitrogenous heterocycle (one six-member pyrimidine ring fused to one five-member imidazole ring) — is the recognition handle that lets ATP-using enzymes bind their substrate with selectivity for the purine over the pyrimidine bases. The ribose sugar is a five-carbon furanose connected to the adenine via a beta-glycosidic bond at N9 of adenine and C1' of ribose; the 2', 3', and 5' hydroxyl groups on ribose participate in further hydrogen bonding and in the phosphate linkage. The triphosphate tail — alpha, beta, and gamma phosphates — is attached at the 5'-hydroxyl of ribose and is the part that does the chemical work.
The adenine moiety contributes essentially nothing to the chemical energetics of ATP hydrolysis itself; the energy lives in the phosphoanhydride bonds (between alpha-beta and beta-gamma phosphates). But the adenine is what makes ATP recognizable as such. Enzymes that use ATP have evolved binding pockets that fit the adenine ring through a stack of aromatic hydrogen bonds and pi-stacking interactions, often involving aspartate or glutamate carboxylates that hydrogen-bond to the N1 and N6 positions of adenine. The "P-loop" motif (GXXXXGKT/S) found in many ATPases is structurally defined by its adenine-binding pocket.
This evolutionary choice is partly arbitrary — biology could have used a pyrimidine-based universal energy currency. But adenine's relatively flat aromatic structure and its hydrogen-bonding pattern are particularly amenable to selective recognition, and once an early life form committed to ATP as energy currency, all subsequent enzyme evolution was constrained to recognize the same molecule. This is why ATP is universal across all three domains of life (bacteria, archaea, eukarya) — the choice was made before the divergence of those lineages roughly 3.5 billion years ago.
The Phosphoanhydride Bond: Where the Energy Lives
The "high-energy" character of ATP is a property of the phosphoanhydride bonds connecting its phosphate groups, not of any single atom. The standard free energy of hydrolysis of ATP→ADP+Pi is approximately -7.3 kcal/mol under standard conditions and closer to -12 kcal/mol under physiological conditions (where ATP is in roughly 10-fold excess over ADP and Pi). For comparison, the hydrolysis of a typical phosphate ester is only about -3 kcal/mol; the phosphoanhydride is about three times more energetic.
Three factors account for the high free energy of phosphoanhydride hydrolysis: resonance stabilization of the released phosphate (the negative charge can delocalize across all four oxygens), electrostatic repulsion within the triphosphate tail at physiological pH (the closely packed negative charges on alpha, beta, and gamma phosphates create unfavorable Coulomb repulsion), and solvation energy changes when the bond breaks. None of these requires unusual bond chemistry — the bonds themselves are normal phosphate-oxygen-phosphate linkages — but the molecular context makes the hydrolysis thermodynamically downhill.
Fritz Lipmann, who first identified the importance of the phosphoanhydride bond in 1941, originally drew the bond with a "squiggle" (~P) to denote its high-energy character. The convention has stuck in biochemistry textbooks ever since, but the squiggle is descriptive shorthand, not a unique bond type.
In cellular terms, the gamma phosphate is the workhorse. Hydrolysis of ATP→ADP+Pi (gamma phosphate removed) is the most common reaction. Less commonly, ATP can be hydrolyzed to AMP + PPi (pyrophosphate), which provides additional driving force because the pyrophosphate is rapidly hydrolyzed by pyrophosphatase to two phosphate groups, making the overall reaction essentially irreversible. This double-hit pattern is used in nucleic acid synthesis (DNA and RNA polymerases) and amino acid activation for protein synthesis (aminoacyl-tRNA synthetases).
ATP Turnover: 60 kg per Day
The cellular ATP pool is small but turns over with astonishing speed. A typical cell contains roughly 1–10 mM ATP, with the whole-body adenine nucleotide pool totaling approximately 250 g (ATP + ADP + AMP combined, mostly in the ATP form). At rest, an adult turns over this pool roughly every 90 seconds.
The daily total ATP turnover for a sedentary 70 kg adult is approximately 60 kg per day — nearly the person's body weight in ATP, regenerated from the same small pool of adenine nucleotides through hundreds of thousands of ADP↔ATP cycles. The corresponding figure for a marathon runner during peak effort is several kilograms per minute. The arithmetic is sometimes cited as one of the most surprising quantities in biology: we run on a tiny inventory of energy molecules, recycled almost as fast as we consume them.
The recycling capacity depends on the integrity of the adenine pool itself. Adenine is rarely destroyed; it is conserved through ADP↔ATP↔AMP interconversion. Only when adenine nucleotides are catabolized to uric acid (via adenosine deaminase → inosine → hypoxanthine → xanthine → uric acid) is the adenine pool depleted. The salvage pathway (APRT + adenine + PRPP → AMP) can reclaim free adenine from blood and tissue breakdown, but if salvage fails (as in APRT deficiency) the pool slowly drains.
This adenine economy explains why purine recycling is so tightly conserved across evolution — cells that wasted adenine through catabolism would be at a metabolic disadvantage. It also explains why mitochondrial disease (failure of oxidative phosphorylation to recycle ADP back to ATP) is so devastating: the issue is not "running out of fuel" in any caloric sense but losing the ability to keep the ATP/ADP ratio high enough to drive endergonic reactions.
Oxidative Phosphorylation and ATP Synthase
The bulk of cellular ATP — roughly 90% under normal aerobic conditions — is synthesized by oxidative phosphorylation in mitochondria. The pathway has two coupled stages.
First, the electron transport chain (Complexes I, II, III, IV embedded in the inner mitochondrial membrane) accepts electrons from NADH and FADH2, passes them down an energy gradient through a series of redox carriers, and ultimately reduces molecular oxygen to water. The energy released at each electron transfer step is used to pump protons from the mitochondrial matrix across the inner membrane into the intermembrane space, building an electrochemical gradient (the "proton motive force") with both a chemical (pH) and an electrical (membrane potential, -180 to -220 mV) component.
Second, ATP synthase (Complex V), a remarkable rotary molecular machine, allows the proton gradient to dissipate by flowing protons back into the matrix through a channel in the F0 subunit. The flow of protons drives the rotation of a central stalk that mechanically forces the F1 catalytic head to cycle through three states — loose, tight, open — binding ADP and Pi in the loose state, condensing them to ATP in the tight state, and releasing ATP in the open state. Each full rotation of the stalk produces three ATP molecules.
The chemiosmotic theory, proposed by Peter Mitchell in 1961 and awarded the Nobel Prize in 1978, was initially controversial because it required a coupling mechanism that was neither a chemical intermediate nor an enzyme cascade but a transmembrane gradient. The eventual structural elucidation of ATP synthase by John Walker (Nobel Prize 1997) confirmed the rotary mechanism in atomic detail.
The adenine nucleotide translocator (ANT) in the inner mitochondrial membrane is the antiporter that exchanges newly synthesized ATP in the matrix for ADP from the cytosol. ANT is one of the most abundant proteins in the inner membrane (10–15% of total inner membrane protein in heart). It is electrogenic: ATP4− out vs ADP3− in, so the exchange is driven by the membrane potential itself. ANT is also the target of the toxin atractyloside, which arrests ATP export and kills cells.
Substrate-Level Phosphorylation in Glycolysis and the TCA Cycle
The remaining 10% of cellular ATP comes from substrate-level phosphorylation in glycolysis and the citric acid cycle. In glycolysis, phosphoglycerate kinase converts 1,3-bisphosphoglycerate + ADP to 3-phosphoglycerate + ATP, and pyruvate kinase converts phosphoenolpyruvate + ADP to pyruvate + ATP. Net glycolytic ATP yield is two per glucose (four made minus two consumed in the priming steps), and these two ATPs are produced regardless of whether oxygen is available — the reason glycolysis can keep cells alive under hypoxia for short periods.
In the citric acid cycle, succinyl-CoA synthetase produces one GTP (or ATP, in some isoforms) per turn. The two pyruvate molecules from each glucose generate two GTP per glucose through this step. GTP and ATP are mutually interconvertible via nucleoside diphosphate kinase, so the cell's adenine and guanine nucleotide pools equilibrate with respect to phosphorylation state.
For rapid bursts of work that exceed oxidative capacity (sprinting, maximal weightlifting), substrate-level phosphorylation cannot keep pace, and the cell relies on the third energy source: the phosphocreatine buffer.
Adenine as Cofactor Scaffold: NAD, FAD, CoA, SAM
Beyond its role in ATP, adenine appears as a structural component in essentially every major redox cofactor and metabolic intermediate carrier.
NAD+ / NADH (nicotinamide adenine dinucleotide) is two nucleotides joined back-to-back through their 5'-phosphates: adenosine monophosphate on one side, nicotinamide mononucleotide on the other. The nicotinamide ring is the actual electron acceptor (cycling between the oxidized NAD+ and reduced NADH forms); the adenine end is the recognition handle. Roughly 400 enzymes in the human proteome use NAD+ as cofactor, including all glycolytic and TCA-cycle dehydrogenases.
NADP+ / NADPH differs from NAD+ only by a single phosphate group esterified to the 2'-hydroxyl of the adenine ribose. That tiny chemical change reroutes the molecule to a different set of enzymes — the reductases of biosynthesis (fatty acid synthesis, cholesterol synthesis, glutathione reduction) rather than the dehydrogenases of catabolism. The cell maintains NADP/NADPH at a much more reduced ratio than NAD/NADH precisely to power reductive biosynthesis.
FAD / FADH2 (flavin adenine dinucleotide) joins adenine to riboflavin (Vitamin B2) through a 5'-5' pyrophosphate linkage. FAD is the cofactor for succinate dehydrogenase (Complex II of the electron transport chain), the fatty acid acyl-CoA dehydrogenases of beta-oxidation, and roughly 80 other enzymes.
Coenzyme A contains an adenine end (attached through pantothenate, Vitamin B5) and a terminal cysteamine thiol that forms thioester bonds with acyl groups. Acetyl-CoA, succinyl-CoA, malonyl-CoA, palmitoyl-CoA — the acyl-thioesters that drive central metabolism — all share the adenine recognition handle. Without it, the acyl-CoA-using enzymes (citrate synthase, acetyl-CoA carboxylase, the beta-oxidation machinery) could not distinguish their substrates from free fatty acids.
S-adenosylmethionine (SAM) is methionine activated by adenosylation at its sulfur, producing a sulfonium center that makes the attached methyl group an excellent leaving group. SAM is the universal methyl donor in biology — involved in DNA methylation (epigenetics), neurotransmitter synthesis (norepinephrine, dopamine), phosphatidylcholine biosynthesis, and roughly 200 other methylation reactions. The byproduct, S-adenosylhomocysteine (SAH), is further metabolized to homocysteine, linking SAM-mediated methylation to folate and B12 status.
The recurring theme is that adenine is biology's "recognition tag" — the molecular barcode that lets cofactor-using enzymes find and bind their substrates with selectivity. The structural conservation across NAD, NADP, FAD, CoA, ATP, SAM, and the cyclic second messengers (cAMP, cGMP) reflects evolutionary convergence on a single recognition motif.
Adenine in Cellular Signaling: cAMP, AMPK, and Adenosine Receptors
Adenine's role extends beyond metabolism into cellular signaling. Three major signaling systems use adenine derivatives directly.
Cyclic AMP (cAMP) is synthesized from ATP by adenylyl cyclase in response to G-protein-coupled receptor activation (epinephrine, glucagon, vasopressin, parathyroid hormone, many others). cAMP is the prototypical second messenger of intracellular signaling, activating protein kinase A (PKA) and the EPAC GTP-exchange factors. cAMP is degraded by phosphodiesterases, with PDE inhibitors (theophylline, caffeine, sildenafil) serving as widely used drugs.
AMP-activated protein kinase (AMPK) is the cellular energy sensor. When ATP is consumed faster than it can be regenerated, the AMP/ATP ratio rises, and AMP binding to AMPK's gamma subunit triggers conformational changes that activate the kinase. Active AMPK then phosphorylates downstream targets to suppress anabolic pathways (lipid synthesis, protein synthesis) and activate catabolic pathways (fatty acid oxidation, autophagy). Metformin, the first-line type 2 diabetes drug, activates AMPK indirectly through mild Complex I inhibition.
Adenosine receptors (A1, A2A, A2B, A3) are G-protein-coupled receptors that respond to extracellular adenosine, which accumulates outside cells when intracellular ATP is depleted (ischemia, exercise, inflammation). A1 receptor activation in the heart slows the AV node — the basis for IV adenosine as a treatment for paroxysmal supraventricular tachycardia. A2A receptors in the basal ganglia regulate dopaminergic transmission — the basis for istradefylline as an adjunctive Parkinson disease drug. A1 receptors in the basal forebrain promote sleep — the basis for caffeine's wakefulness effect (caffeine is a competitive adenosine receptor antagonist).
The fact that adenine derivatives signal in all three intracellular regimes (ATP as substrate, cAMP as second messenger, AMP as energy sensor) plus the extracellular regime (adenosine on cell-surface receptors) reflects deep evolutionary co-option. The same molecular scaffold serves as energy currency, signal carrier, and danger sign — one molecule wearing four hats.
Muscle Energetics: Creatine Phosphate Buffer
Skeletal muscle and heart have an additional energy storage system that buffers the ATP pool against sudden demand: phosphocreatine (also called creatine phosphate). Resting muscle maintains a phosphocreatine concentration roughly four times higher than ATP. When ATP is consumed during contraction, creatine kinase rapidly transfers the high-energy phosphate from phosphocreatine to ADP, regenerating ATP within milliseconds.
The reaction (PCr + ADP ↔ Cr + ATP) is freely reversible and equilibrates in microseconds. During sprinting, phosphocreatine is depleted within 8–10 seconds, after which the muscle must rely on glycolysis and (later) oxidative phosphorylation. During recovery, oxidative phosphorylation regenerates the phosphocreatine pool by reversing the reaction.
This is why creatine supplementation (typically 3–5 g/day of monohydrate) is one of the few supplements with robust evidence for short-burst athletic performance — it increases muscle phosphocreatine stores by about 20%, expanding the immediate ATP buffer.
The adenine connection: creatine kinase recognizes ADP and ATP through their adenine moieties, so the entire phosphocreatine buffer system depends on the adenine recognition motif evolved for the broader ATP economy.
Mitochondrial Disease and ATP Production Failure
When the ATP synthesis machinery fails, the clinical consequences are predictably severe. Mitochondrial diseases — caused by mutations in nuclear-encoded or mitochondrially-encoded subunits of the electron transport chain or ATP synthase, or by defects in mitochondrial DNA replication and maintenance — preferentially affect tissues with high ATP demand: brain, heart, skeletal muscle, and the retina.
Common clinical syndromes include MELAS (mitochondrial encephalomyopathy with lactic acidosis and stroke-like episodes), MERRF (myoclonic epilepsy with ragged-red fibers), LHON (Leber hereditary optic neuropathy), Kearns-Sayre syndrome, Pearson syndrome, NARP (neuropathy, ataxia, retinitis pigmentosa), and Leigh syndrome. The molecular common denominator is impaired ATP synthesis, with lactic acidosis as a near-universal biochemical feature (reflecting the cell's switch to anaerobic glycolysis when oxidative phosphorylation fails).
Therapies for mitochondrial disease remain limited. Coenzyme Q10 supplementation (a component of the electron transport chain), idebenone (a synthetic CoQ10 analog), riboflavin (which supports Complex II via FAD), thiamine (which supports the pyruvate dehydrogenase complex), and L-carnitine (which supports fatty acid import into mitochondria) are commonly used but provide modest benefit. Gene therapy approaches (allotopic expression, mitochondrial replacement therapy) remain experimental.
The relevance to adenine: defective oxidative phosphorylation does not deplete the adenine pool itself (the body conserves adenine through salvage), but it cripples the cell's ability to keep that pool in the ATP form. Cellular ATP/ADP ratios crash, AMP rises, AMPK is chronically activated, and the cell shifts to a catabolic state that, sustained over months and years, manifests as the progressive multi-organ failure of mitochondrial disease.
Key Research Papers
- Lipmann F, generation and utilization of high-energy phosphate bonds (1941) — PubMed: Lipmann 1941
- Mitchell P, chemiosmotic coupling of oxidative phosphorylation (1961 Nature) — PubMed: Mitchell 1961
- Boyer PD, binding-change mechanism of ATP synthase (Nobel) — PubMed: Boyer binding-change
- Walker JE, structural elucidation of F1 ATP synthase (1994 Nature) — PubMed: Walker F1 structure
- Pebay-Peyroula E, structure of the adenine nucleotide translocator ANT — PubMed: ANT structure
- Hardie DG, AMP-activated protein kinase as cellular energy sensor — PubMed: Hardie AMPK
- Sutherland EW, discovery of cyclic AMP and second messenger concept — PubMed: Sutherland cAMP
- Fredholm BB, adenosine receptor pharmacology (A1, A2A, A2B, A3) — PubMed: Fredholm receptors
- Wallimann T, creatine kinase circuit and phosphocreatine buffer in muscle — PubMed: Wallimann CK circuit
- DiMauro S, mitochondrial respiratory chain diseases review — PubMed: DiMauro mitochondrial disease
- Wallace DC, mitochondrial DNA in human aging and disease — PubMed: Wallace mtDNA
- Imai S, NAD+ and sirtuin biology in aging — PubMed: Imai NAD & sirtuins
Connections
- Vitamin B4 Benefits (Hub)
- Vitamin B4 (Main Page)
- Purine Metabolism
- DNA and RNA Synthesis
- Modern Status and Food Sources
- Vitamin B2 (Riboflavin / FAD)
- Vitamin B3 (Niacin / NAD)
- Vitamin B5 (Pantothenic Acid / CoA)
- Magnesium (ATP cofactor)
- Phosphorus
- Creatine
- Coenzyme Q10
- Mitochondrial Disease
- Type 2 Diabetes (AMPK / Metformin)
- All Vitamins