Copper for Connective Tissue
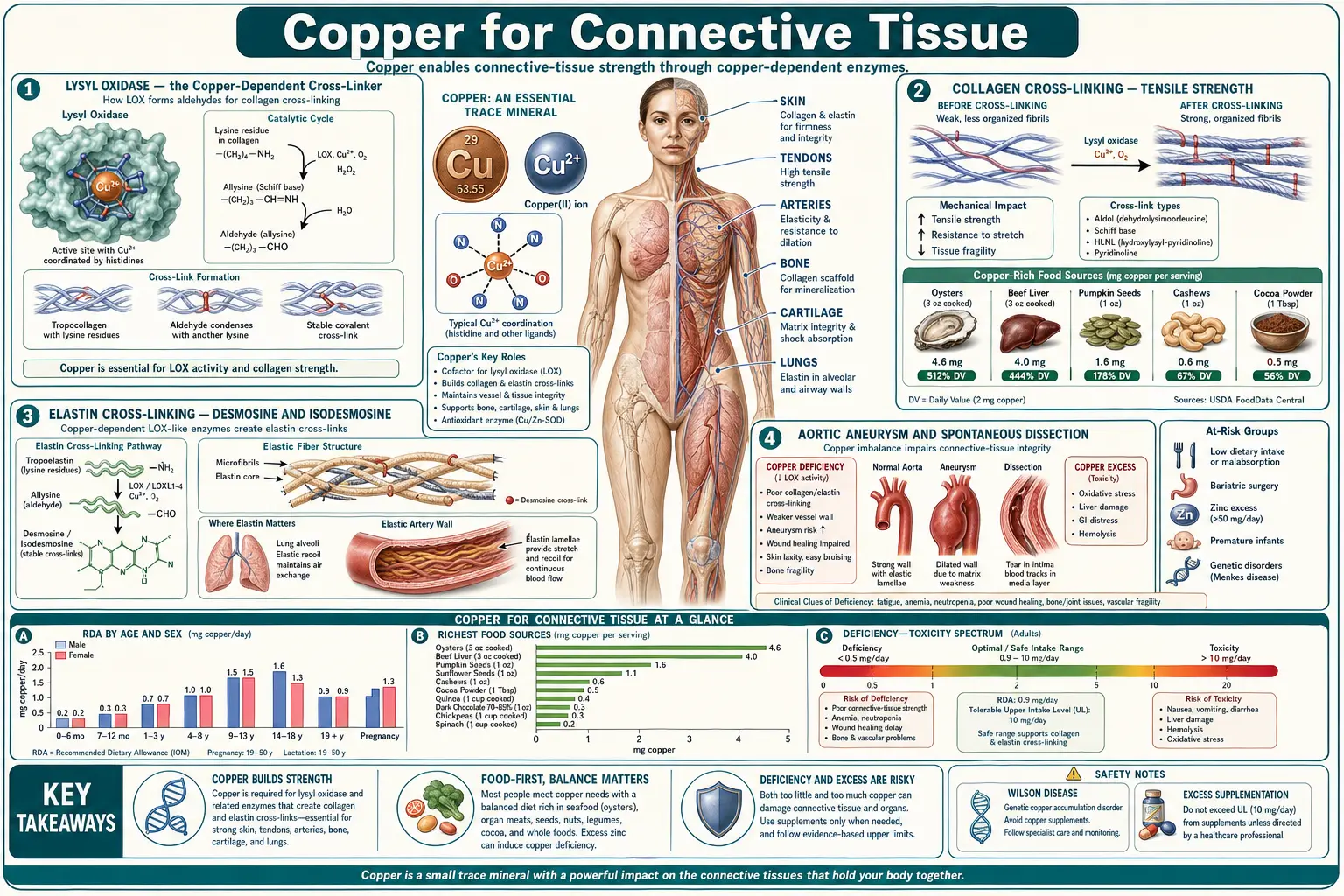
Copper is the catalytic atom at the active site of lysyl oxidase (LOX), the extracellular enzyme that creates the covalent cross-links holding collagen fibers and elastin sheets together. Without lysyl oxidase activity, tropocollagen monomers cannot polymerize into the staggered, cross-linked fibrils that give skin tensile strength and tendons their pull-resistance, and tropoelastin monomers cannot form the desmosine and isodesmosine ring structures that give the aortic wall and lung parenchyma their elastic recoil. The clinical consequences of severe copper deficiency — whether genetic (Menkes disease, occipital horn syndrome) or acquired (zinc overdose, bariatric surgery, prolonged TPN) — therefore include aortic aneurysm and dissection, fragile bones with reduced cortical thickness, "kinky" or twisted hair (pili torti), loose redundant skin, bladder diverticula, and joint hypermobility. The vascular fragility is the most clinically lethal manifestation. This page walks through the lysyl oxidase mechanism, the cross-linking chemistry, the genetic and acquired syndromes that demonstrate the requirement, and the practical implications for cardiovascular and orthopedic health.
Table of Contents
- Lysyl Oxidase — the Copper-Dependent Cross-Linker
- Collagen Cross-Linking — Tensile Strength
- Elastin Cross-Linking — Desmosine and Isodesmosine
- Aortic Aneurysm and Spontaneous Dissection
- Menkes Disease — the Genetic Proof
- Occipital Horn Syndrome (Mild Menkes)
- Bone Fragility and Cortical Thinning
- Skin Elasticity, Pili Torti, and Wound Healing
- Acquired Vascular Risk in Adults
- Topical Copper Peptides (GHK-Cu) for Skin
- Key Research Papers
- Connections
- Featured Videos
Lysyl Oxidase — the Copper-Dependent Cross-Linker
Lysyl oxidase (LOX, EC 1.4.3.13) is a 32-kilodalton extracellular copper-containing enzyme that catalyzes the oxidative deamination of peptidyl lysine and hydroxylysine residues in collagen and elastin. The reaction is:
peptidyl-lysine + O&sub2; + H&sub2;O → peptidyl-allysine + NH&sub3; + H&sub2;O&sub2;
The peptidyl-allysine product is a reactive aldehyde that spontaneously condenses with other allysines (or with unmodified lysines) on adjacent collagen or elastin chains to form Schiff-base and aldol cross-links. These cross-links are what convert the soluble monomeric precursors (tropocollagen, tropoelastin) into the insoluble cross-linked fibrils and fibers that give connective tissue its mechanical properties.
LOX is synthesized as a 50-kDa proenzyme by fibroblasts, smooth muscle cells, chondrocytes, and other matrix-producing cells. It is secreted into the extracellular space and proteolytically activated by bone morphogenetic protein 1 (BMP-1), which removes an N-terminal propeptide to release the active 32-kDa mature enzyme. The mature enzyme contains one copper atom per molecule plus a covalently bound cofactor called lysyl tyrosylquinone (LTQ), which is formed by intramolecular cross-linking of a copper-coordinated lysine and a tyrosine residue in the enzyme itself. Both the copper atom and the LTQ cofactor are essential for catalysis — remove either and the enzyme is inactive.
The copper-loading step happens in the late secretory pathway through the copper-transporting ATPase ATP7A. Loss-of-function mutations in ATP7A — the cause of Menkes disease — therefore produce a phenotype that includes severe lysyl oxidase deficiency on top of the systemic copper deficiency, because the enzyme is synthesized and secreted as the inactive copper-free apoprotein.
There are five LOX-family members in humans: LOX itself plus LOXL1, LOXL2, LOXL3, and LOXL4. Each performs similar oxidative chemistry on different connective-tissue substrates in different tissues, and all are copper-dependent. LOXL1 deficiency is associated with pelvic organ prolapse and pseudoexfoliation glaucoma; LOX deficiency is the dominant contributor to aortic and skin pathology.
Collagen Cross-Linking — Tensile Strength
Collagen is the most abundant protein in the body (approximately 30% of total body protein) and the primary tensile-load-bearing protein in skin, bone, tendon, ligament, cartilage, and the dermis. It is synthesized as procollagen by fibroblasts, secreted into the extracellular space, proteolytically processed to tropocollagen, and assembled into the staggered quarter-stagger array that produces collagen fibrils with their characteristic 67-nanometer banding pattern. The fibrils gain their tensile strength from intermolecular covalent cross-links introduced by lysyl oxidase.
The cross-linking chemistry begins with LOX oxidation of specific lysine and hydroxylysine residues in the N- and C-terminal telopeptide regions of tropocollagen. The resulting aldehydes (allysine and hydroxyallysine) then participate in spontaneous, non-enzymatic condensation reactions to form first immature divalent cross-links (such as hydroxylysinonorleucine and dehydrohydroxylysinonorleucine), then mature trivalent cross-links (pyridinoline and deoxypyridinoline) over weeks to months. The mature trivalent cross-links are the most mechanically important and the slowest to form.
Without lysyl oxidase, this cross-linking does not occur. The tropocollagen monomers do assemble into fibrils, but they are mechanically weak. Skin in copper deficiency becomes thinner, more easily torn, and slower to heal. Tendons and ligaments lose tensile strength. The wound-healing defect is particularly important in surgical and burn patients with marginal copper status.
Elastin Cross-Linking — Desmosine and Isodesmosine
Elastin is the second great fibrous protein of the connective tissue and the one that gives the aorta, lung, skin, and ligamentum flavum their elastic recoil. Where collagen resists tension, elastin stretches and then recoils elastically — the aortic wall, for example, must expand by about 10% with each systolic ejection and recoil during diastole, billions of times over a lifetime, without fatigue or failure.
The elastic-recoil property comes from elastin's unusual cross-linking architecture. Tropoelastin monomers are coiled into a hydrophobic, water-resistant rope-like structure and then cross-linked by lysyl oxidase. But where collagen cross-linking uses simple divalent or trivalent bonds, elastin uses tetravalent cross-links — desmosine and isodesmosine, which are pyridinium ring structures formed by the condensation of four lysine residues (three of which have been converted to allysine by LOX) on adjacent tropoelastin chains.
Desmosine and isodesmosine are unique to elastin — no other protein contains them. The tetravalent cross-linking creates a covalently bonded three-dimensional network that allows elastin to stretch and recoil without breaking or unraveling. The cross-link density is extremely high in tissues that must endure repeated mechanical strain: the aortic media, the elastic ligaments, and the elastic fibers of the lung.
Copper deficiency reduces lysyl oxidase activity and therefore reduces desmosine and isodesmosine formation. The result is "weakened" elastin — mechanically inadequate for the cyclic loading that healthy aortic wall and lung parenchyma must endure. Animal studies in copper-deficient pigs, rats, and chickens consistently show reduced elastin desmosine content along with arterial wall thinning, aneurysm formation, and skin laxity.
Aortic Aneurysm and Spontaneous Dissection
The most clinically dangerous manifestation of copper deficiency is failure of the aortic wall. The thoracic aorta, in particular, depends on intact elastin for its ability to absorb the systolic pulse pressure and recoil during diastole. When elastin cross-linking is inadequate, the aortic wall undergoes medial degeneration — loss of elastic lamellae, replacement by amorphous extracellular matrix, smooth muscle cell dropout, and ultimately frank dilation (aneurysm) or tearing (dissection).
The classic experimental demonstration came from studies of copper-deficient pigs in the 1960s and 1970s — the animals developed spontaneous aortic aneurysm and rupture, a phenotype that resolved with copper repletion. Similar work in copper-deficient chickens (Tinker and Rucker 1985) established that aortic elastin desmosine content fell by 50–70% with severe copper deficiency, paralleled by reduced aortic tensile strength.
In humans, the genetic conditions that disrupt copper-dependent lysyl oxidase activity — Menkes disease and occipital horn syndrome (see below) — produce aortic aneurysm and arterial tortuosity as core features. Beyond these rare genetic conditions, acquired copper deficiency in adults has been implicated in some thoracic aortic aneurysm cases, though the evidence is more circumstantial than for the genetic conditions. The Lox knockout mouse (genetic deletion of LOX) dies perinatally from aortic and diaphragmatic rupture, providing rigorous proof that LOX activity is essential for vascular integrity.
The practical clinical implication: patients with the acquired copper-deficiency risk factors (zinc overdose, bariatric surgery, prolonged TPN, severe malabsorption) and especially those with first-degree family history of thoracic aortic aneurysm, dissection, or sudden cardiac death should have copper status assessed. The cardiac surgery and vascular medicine literature is still catching up to the recognition that nutritional copper deficiency is a real (though under-screened) modifiable risk factor for some aortic disease.
Menkes Disease — the Genetic Proof
Menkes disease (also called Menkes kinky hair syndrome) is the X-linked recessive disorder caused by loss-of-function mutations in ATP7A, the copper-transporting ATPase on the basolateral membrane of the enterocyte. ATP7A is required to export dietary copper from the enterocyte into the portal circulation, and a parallel ATP7A pool in the late secretory pathway of all cells delivers copper to the metalation step of cuproenzymes including lysyl oxidase, dopamine beta-hydroxylase, and tyrosinase.
Loss of ATP7A produces systemic copper deficiency from infancy — the affected boy (Menkes is X-linked, so virtually all clinically affected patients are male) absorbs copper into the enterocyte but cannot export it. Dietary copper accumulates in the gut wall and is shed with the enterocyte at the end of its lifespan, never reaching the rest of the body. The clinical phenotype emerges in the first months of life:
- "Kinky" or twisted hair (pili torti) — the pathognomonic finding, with hair that is sparse, lusterless, brittle, and microscopically shows twisting along the long axis. The disrupted hair cross-linking is the visible manifestation of LOX deficiency.
- Intractable seizures and developmental regression — from copper-dependent enzyme failures in the developing brain
- Hypothermia — from impaired norepinephrine synthesis (dopamine beta-hydroxylase is copper-dependent)
- Hypotonia and feeding difficulties
- Pale skin — from tyrosinase failure (tyrosinase is copper-dependent and is the rate-limiting enzyme of melanin synthesis)
- Aortic dilation, arterial tortuosity, and intracranial vascular abnormalities — the LOX-deficiency vascular phenotype
- Bladder diverticula and inguinal hernia — from extracellular matrix weakness
- Bone abnormalities — "Wormian bones" in the skull, metaphyseal flaring, and fragility
- Death typically by 3 years of age in untreated classical Menkes disease, usually from infection or vascular catastrophe
Early treatment with subcutaneous copper-histidine injections (started in the first weeks of life, before significant brain copper deficiency) can extend survival and partially preserve neurologic function in some milder ATP7A variants, but does not fully rescue the phenotype in classical Menkes. The disease provides the cleanest in vivo demonstration of what happens when all copper-dependent enzymes fail simultaneously.
Occipital Horn Syndrome (Mild Menkes)
Occipital horn syndrome (OHS) is the milder allelic variant of Menkes disease, caused by ATP7A mutations that retain partial function (typically 5–30% of normal activity). Where classical Menkes is lethal in early childhood, OHS patients survive into adulthood with a connective-tissue-dominant phenotype:
- Occipital exostoses (the "horns") — bilateral bony protrusions from the occipital bone, the eponymous feature, detectable on plain skull radiograph
- Skin laxity — loose, soft skin, particularly on the neck and trunk
- Joint hypermobility — not Marfan-syndrome severity but clinically apparent
- Bladder diverticula — from extracellular matrix weakness in the bladder wall
- Inguinal and umbilical hernias
- Mild cognitive impairment — less severe than classical Menkes
- Arterial tortuosity — visible on angiography
OHS was historically classified as "Ehlers-Danlos syndrome type IX" before its molecular cause was identified, reflecting how directly the phenotype mimics other heritable connective-tissue disorders. The current Villefranche EDS nomenclature has removed type IX, recognizing OHS as a copper-deficiency disorder rather than a primary collagen disorder. The pathophysiologic distinction is moot for clinical purposes — LOX deficiency produces a phenotype that overlaps with collagen-mutation EDS because LOX is the cross-linking enzyme that finishes the collagen story.
Bone Fragility and Cortical Thinning
Copper deficiency produces a characteristic bone phenotype: reduced cortical thickness, metaphyseal flaring, "rib-end" beading mimicking rickets or scurvy, increased fragility, and reduced bone density on DEXA. The mechanism is again primarily lysyl oxidase — the collagen scaffold of bone is the framework on which hydroxyapatite mineral is deposited, and inadequate collagen cross-linking produces a mechanically weaker bone even when mineralization is otherwise normal.
The classic clinical setting is the copper-deficient infant on cow-milk formula (Cordano A et al. 1964) or the long-term TPN patient with inadequate copper. Both groups show characteristic radiographic findings: anterior rib flaring, metaphyseal cupping, osteoporosis with fractures, and "wormian bones" (extra ossification centers in the cranial sutures).
The adult copper-deficiency bone phenotype is less dramatic but still consequential. Population studies suggest a modest association between marginal copper status and reduced bone mineral density and increased fracture risk, particularly in elderly women. The clinical bottom line: in any patient with unexplained osteoporosis, fracture beyond what bone density would predict, or family history of fragility fracture, copper status is worth assessing alongside the conventional Vitamin D, calcium, and PTH workup.
Skin Elasticity, Pili Torti, and Wound Healing
The skin is the most accessible tissue for copper-deficiency assessment, and the hair is among the most sensitive markers. Pili torti — hair shafts twisted around their longitudinal axis, visible on dermoscopy — is the pathognomonic finding of Menkes disease and is also seen in severe acquired copper deficiency. Hair lightening (from tyrosinase failure) is another finding, more subtle in adults than in the strikingly hypopigmented Menkes infant.
Skin laxity, easy bruising, fragile capillaries, and slow wound healing are common in copper deficiency. The mechanism is the same: inadequate collagen and elastin cross-linking produces a dermis that does not maintain its shape under load and does not heal effectively after injury.
The wound-healing application has clinical significance. Surgical patients, burn patients, and patients with chronic wounds (diabetic foot ulcers, pressure ulcers) are at risk if their copper status is marginal. Copper supplementation as part of nutritional support in these settings has demonstrated benefit in observational and small randomized series, though it is not yet standard of care in most wound clinics.
For broader skin-health interventions, see our pages on Acne, Collagen, and the related antioxidant Vitamin C (which is the cofactor for prolyl and lysyl hydroxylase, the enzymes that hydroxylate the collagen precursor before LOX cross-links it — meaning Vitamin C and copper act in series on the same collagen molecules).
Acquired Vascular Risk in Adults
Beyond the rare genetic conditions, acquired copper deficiency in adults is increasingly recognized as a contributor to vascular pathology. The high-risk populations are:
- Post-bariatric surgery patients — particularly Roux-en-Y gastric bypass and duodenal switch, who bypass the major copper-absorption region of the duodenum and proximal jejunum. The published literature includes case reports of post-bariatric patients presenting with thoracic aortic aneurysm or dissection in the setting of profound copper deficiency, alongside the more familiar copper-deficiency myelopathy.
- Long-term zinc overdose — chronic zinc supplementation above 50 mg/day, denture cream with high zinc content (legacy formulations), or zinc-rich occupational exposure (rare).
- Severe malabsorption — refractory celiac, advanced Crohn's, short-bowel syndrome, chronic pancreatic insufficiency.
- Long-term parenteral nutrition — with inadequate trace mineral supplementation.
- Wilson disease patients on aggressive zinc or chelation therapy — iatrogenic copper deficiency if monitoring lapses.
The clinical recommendation in these populations is to monitor copper, ceruloplasmin, and zinc status periodically (typically every 6–12 months), and to maintain copper status in the lower-normal to mid-normal range rather than the lowest-acceptable end. The cost of supplementation is trivial; the cost of a missed copper deficiency leading to vascular catastrophe is enormous.
Topical Copper Peptides (GHK-Cu) for Skin
The tripeptide glycyl-L-histidyl-L-lysine (GHK), when complexed with copper as GHK-Cu, has a long history in topical dermatology and wound-care formulations. GHK occurs naturally in human plasma and is thought to act as a copper carrier and a tissue-remodeling signaling peptide. Topical GHK-Cu products have been marketed for anti-aging skincare since the 1980s, and the published in vitro and animal-model literature supports several mechanisms:
- Stimulates collagen and elastin synthesis in cultured fibroblasts
- Activates lysyl oxidase by delivering copper to the active site
- Increases proteoglycan and glycosaminoglycan synthesis in the dermis
- Modulates matrix metalloproteinase activity, favoring tissue remodeling over degradation
- Improves wound healing in animal models of diabetic and pressure ulcers
The human clinical evidence is more limited — small open-label and short-term randomized cosmetic studies of topical GHK-Cu serums show modest improvement in skin firmness, fine wrinkles, and elasticity scores, but rigorous large randomized trials are scarce. The mechanistic basis is plausible (delivering copper transdermally to the dermal fibroblasts that synthesize lysyl oxidase), and the safety profile is excellent. GHK-Cu is best considered an adjunctive approach to skin health rather than a substitute for adequate systemic copper status from diet.
Key Research Papers
- Kagan HM, Trackman PC (1991). Properties and function of lysyl oxidase. American Journal of Respiratory Cell and Molecular Biology. — PubMed
- Tinker D, Rucker RB (1985). Role of selected nutrients in synthesis, accumulation, and chemical modification of connective tissue proteins. Physiological Reviews. — PubMed
- Máki JM et al. (2002). Inactivation of the lysyl oxidase gene Lox leads to aortic aneurysms, cardiovascular dysfunction, and perinatal death in mice. Circulation. — PubMed
- Kaler SG (2011). ATP7A-related copper transport diseases — emerging concepts and future trends. Nature Reviews Neurology. — PubMed
- Cordano A, Baertl JM, Graham GG (1964). Copper deficiency in infancy. Pediatrics. — PubMed
- Mercer JF (1998). Menkes syndrome and animal models. American Journal of Clinical Nutrition. — PubMed
- Rucker RB et al. (1998). Copper, lysyl oxidase, and extracellular matrix protein cross-linking. American Journal of Clinical Nutrition. — PubMed
- Pickart L, Margolina A (2018). Regenerative and protective actions of the GHK-Cu peptide in the light of the new gene data. International Journal of Molecular Sciences. — PubMed
- Tang S et al. (1989). Reduced lysyl oxidase activity and arterial wall connective tissue in copper-deficient swine. Journal of Nutrition. — PubMed
- Liu G et al. (2004). A genetic model of CADASIL revealed in lysyl oxidase deficient mice. Journal of Investigative Dermatology. — PubMed
- Strecker MN et al. (2002). Occipital horn syndrome: clinical phenotype and natural history. American Journal of Medical Genetics. — PubMed
- Tinker D, Geller J, Romero N, Wedge JR, Rucker RB (1990). Cardiovascular and connective tissue effects of dietary copper depletion in rats. Journal of Nutrition. — PubMed
PubMed Topic Searches
- PubMed: Lysyl oxidase copper cross-linking
- PubMed: Copper deficiency and aortic aneurysm
- PubMed: Menkes disease
- PubMed: Occipital horn syndrome
- PubMed: Copper and bone fragility
Connections
- Copper Overview
- Copper Benefits Hub
- Beef Liver for Copper & Iron
- Copper for Hemoglobin and Ceruloplasmin
- Copper for Antioxidant Defense
- Copper for Neurological Health
- Zinc (Copper Antagonist)
- Magnesium
- Vitamin C (Collagen Hydroxylation Cofactor)
- Vitamin A
- Collagen
- Organ Meats (Beef Liver)
- Morley Robbins
- Whole Food Copper Sources
- Cardiology
- Dermatology
- Beef
- Dark Chocolate