Japanese Knotweed Anti-Inflammatory & Antioxidant Effects
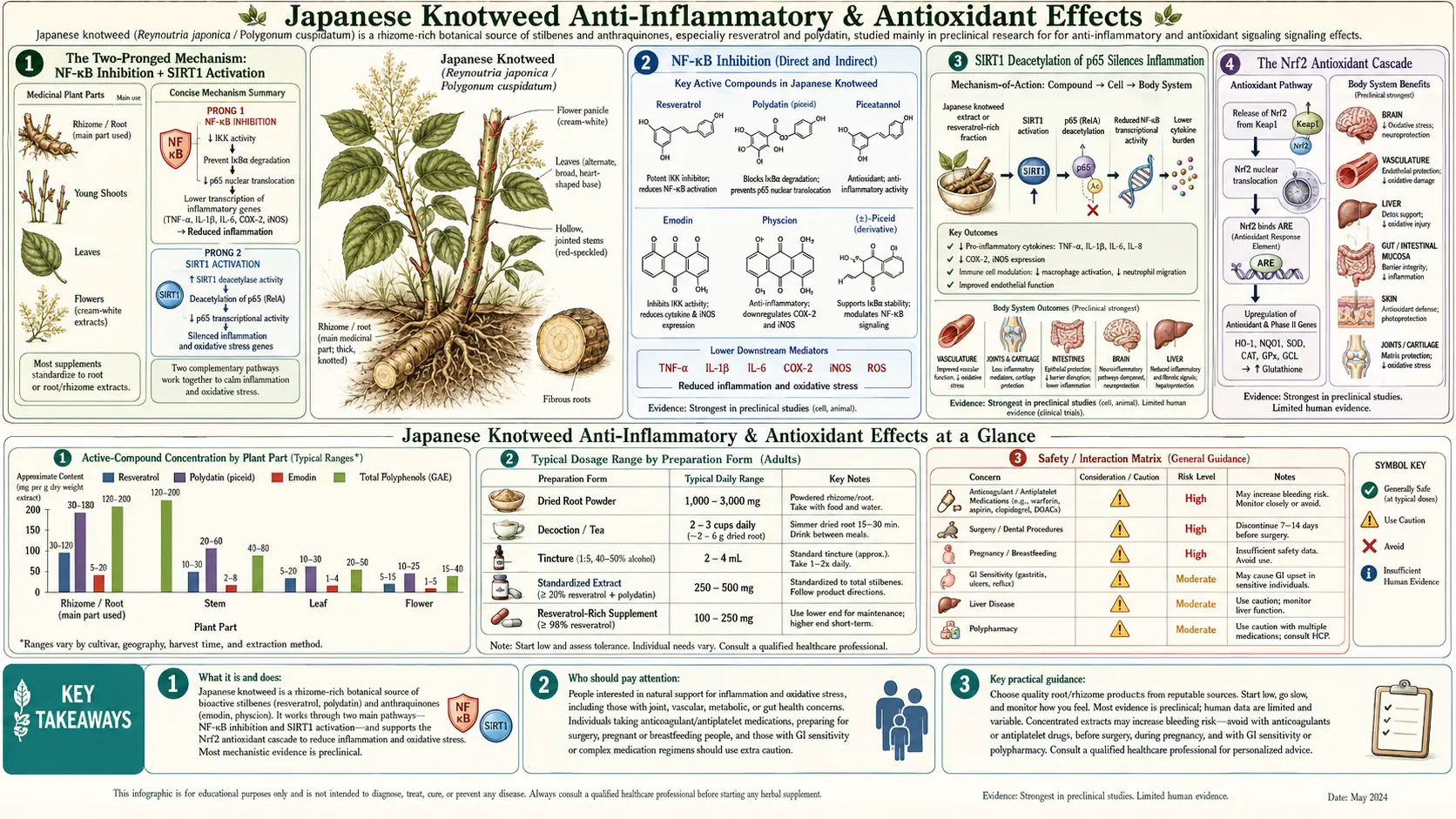
Japanese knotweed produces its anti-inflammatory effects through a two-pronged molecular mechanism that explains why the herb works across such a wide range of inflammatory conditions: resveratrol inhibits NF-κB (the master transcription factor for inflammatory gene expression) AND activates SIRT1 (the NAD+-dependent deacetylase that removes acetyl groups from the NF-κB p65 subunit to silence its transcriptional activity). These two mechanisms reinforce each other — one prevents NF-κB activation in the first place, the other shuts down NF-κB activity that has already started. A third pathway, Nrf2 activation, raises the body's endogenous antioxidant capacity rather than supplying antioxidants directly. Emodin, the anthraquinone constituent of knotweed root, adds independent anti-inflammatory and antifungal effects. Traditional Chinese Medicine recognized this multi-mechanism profile centuries ago, classifying knotweed as a herb for "blood stasis" — the TCM term for the combination of inflammation, oxidation, and impaired circulation that drives so much chronic disease.
Table of Contents
- The Two-Pronged Mechanism: NF-κB Inhibition + SIRT1 Activation
- NF-κB Inhibition (Direct and Indirect)
- SIRT1 Deacetylation of p65 Silences Inflammation
- The Nrf2 Antioxidant Cascade
- COX-2 and iNOS Downregulation
- Emodin (Anthraquinone) Effects
- Traditional Chinese Medicine: "Blood Stasis" and Inflammation
- Arthritis and Joint Inflammation
- Inflammatory Bowel Disease
- Skin Inflammation and Atopic Dermatitis
- Neuroinflammation and Microglial Modulation
- Oxidative Stress in Chronic Disease
- Cautions
- Key Research Papers
- Connections
- Featured Videos
The Two-Pronged Mechanism: NF-κB Inhibition + SIRT1 Activation
The central insight that explains Japanese knotweed's anti-inflammatory profile is that resveratrol acts on the inflammatory cascade through two complementary mechanisms that work in different directions on the same target. Most anti-inflammatory drugs hit a single mechanism: aspirin and NSAIDs inhibit COX enzymes, corticosteroids bind glucocorticoid receptors and broadly suppress gene expression, biological agents (anti-TNF antibodies, IL-6 receptor antagonists) neutralize individual cytokines. Resveratrol hits the master inflammatory transcription factor NF-κB both upstream (preventing activation) and downstream (silencing activated p65) of its activation cycle.
Understanding this requires a brief tour of the NF-κB pathway. NF-κB is a dimeric transcription factor (usually p65/p50 in classical inflammation) that sits in the cytoplasm bound to its inhibitor IκB. When the cell receives an inflammatory signal (TLR engagement by bacterial products, TNF receptor engagement, IL-1 receptor engagement, oxidative stress, etc.), a kinase cascade phosphorylates IκB at specific serines, marking it for ubiquitination and proteasomal degradation. Free of its inhibitor, NF-κB translocates to the nucleus, where it binds κB response elements in the promoters of hundreds of target genes — COX-2, iNOS, TNF-α, IL-1β, IL-6, IL-8, MMP-9, ICAM-1, VCAM-1, E-selectin, and many more. The p65 subunit gets acetylated at lysines 218, 221, and 310 by the p300/CBP acetyltransferase; acetylation is required for full transcriptional activity. The transcriptional response continues until p65 is deacetylated and re-exported to the cytoplasm, where new IκB (also a NF-κB target gene) re-binds it.
Resveratrol's two mechanisms hit this cycle at different points:
- Upstream (NF-κB inhibition) — resveratrol interferes with the IκB kinase (IKK) complex, preventing efficient phosphorylation of IκB. The result is reduced NF-κB activation in response to inflammatory stimuli. This effect is direct and dose-dependent.
- Downstream (SIRT1-mediated p65 deacetylation) — resveratrol activates SIRT1, which deacetylates p65 at lysine 310. Deacetylation reduces p65's ability to bind κB response elements and recruit transcriptional machinery. This effect silences NF-κB activity that has already been initiated.
The combination produces a more thorough silencing of inflammatory gene expression than either mechanism alone, which is why resveratrol-rich extracts can produce measurable cytokine suppression even at modest tissue concentrations. The mechanism also explains why the anti-inflammatory effect is observed across such a wide range of conditions — the central transcription factor of inflammation is the same regardless of the upstream trigger.
NF-κB Inhibition (Direct and Indirect)
NF-κB inhibition by resveratrol has been documented in multiple cell types and inflammatory contexts. The mechanism involves several molecular events:
- Direct IKK inhibition — resveratrol binds to the IKKβ subunit of the IκB kinase complex, reducing its ability to phosphorylate IκB
- Reduced IκB degradation — as a consequence of reduced phosphorylation, IκB remains bound to NF-κB and the dimer remains in the cytoplasm
- Reduced nuclear translocation — the small fraction of NF-κB that does activate translocates less efficiently to the nucleus
- Reduced DNA binding — nuclear NF-κB binds κB response elements with lower affinity in the presence of resveratrol
- Cross-talk with other inflammatory pathways — reduced NF-κB activity propagates to reduced MAPK signaling (p38, JNK, ERK) and reduced STAT3 activity, broadening the anti-inflammatory effect
The net effect on inflammatory gene expression is a 30 to 60% reduction in NF-κB target gene transcription in response to a given inflammatory stimulus, depending on cell type, stimulus, and resveratrol concentration. This is a substantial molecular-level effect that translates to measurable cytokine suppression in vivo.
One important nuance: NF-κB inhibition is not the same as "turning off inflammation." NF-κB has essential roles in immunity against infection, in tissue repair, in development, and in cell survival. Complete NF-κB inhibition would be catastrophic; what resveratrol produces is a modulation — turning down the volume on excessive or chronic inflammation while leaving acute and necessary inflammation largely intact. This is part of why resveratrol does not produce the immune suppression and infection susceptibility associated with corticosteroids and biological anti-cytokine therapy.
SIRT1 Deacetylation of p65 Silences Inflammation
The SIRT1 arm of the anti-inflammatory mechanism is mechanistically distinct from the IKK arm. Even when NF-κB has been activated (whether by stimuli too strong for IKK inhibition to fully prevent, or by stimuli that bypass IKK), the activity of nuclear NF-κB depends on the acetylation status of its p65 subunit. SIRT1 reverses this acetylation, providing a downstream brake on inflammatory transcription.
The key acetylation site on p65 is lysine 310. Acetyl-K310 is required for full transcriptional activity at NF-κB target genes, particularly for the recruitment of the p300/CBP coactivator complex that bridges NF-κB to the basal transcriptional machinery. SIRT1 specifically deacetylates K310 (it can deacetylate other p65 lysines as well, but K310 is the primary functional target). Deacetylated p65 either fails to recruit coactivators or recruits them less efficiently, silencing transcription of the bound target genes.
This pathway is part of a broader principle: SIRT1 acts as a metabolic-state-responsive brake on inflammation. Under conditions of low cellular energy (caloric restriction, fasting, sirtuin pathway activation), high NAD+/NADH ratios drive SIRT1 activity, which suppresses excessive inflammatory responses that the body cannot afford energetically. Under conditions of high cellular energy (overnutrition, sedentariness, low NAD+), SIRT1 activity falls, removing this brake and allowing the chronic low-grade inflammation that drives metabolic and age-related disease. Resveratrol works partly by pharmacologically restoring SIRT1 activity that has been lost to age-related NAD+ decline.
The clinical translation: chronic low-grade inflammation ("inflammaging") is a hallmark of aging and a driver of most age-related diseases. Restoring SIRT1 activity through pharmacologic activation (resveratrol) or NAD+ supplementation (NR, NMN) is the unifying intervention proposed by the longevity research field. The anti-inflammatory effects observed in resveratrol trials are partly explained by this mechanism.
The Nrf2 Antioxidant Cascade
The third pillar of resveratrol's anti-inflammatory/antioxidant effect is activation of the nuclear factor erythroid 2-related factor 2 (Nrf2) transcription factor. Nrf2 is the master regulator of the cellular antioxidant defense system. Under basal conditions, Nrf2 is held in the cytoplasm by its inhibitor Keap1 (Kelch-like ECH-associated protein 1), which targets Nrf2 for proteasomal degradation. Under oxidative stress (or in response to specific Keap1-modifying compounds like resveratrol), Keap1 releases Nrf2, which translocates to the nucleus and binds the antioxidant response element (ARE) in the promoter regions of dozens of antioxidant and phase II detoxification genes.
The Nrf2 target gene battery includes:
- Glutamate-cysteine ligase (GCL) — the rate-limiting enzyme for glutathione synthesis
- Glutathione S-transferases (GSTs) — conjugate glutathione to electrophilic toxins for excretion
- NAD(P)H quinone dehydrogenase 1 (NQO1) — detoxifies quinones and prevents their redox cycling
- Heme oxygenase-1 (HO-1) — degrades heme to bilirubin (a potent antioxidant), CO (a signaling molecule), and iron (sequestered by ferritin)
- Thioredoxin and thioredoxin reductase — major cellular reducing system
- Superoxide dismutase (SOD) — converts superoxide to hydrogen peroxide
- Catalase — converts hydrogen peroxide to water
- Glutathione peroxidases (GPx) — convert hydrogen peroxide and lipid peroxides to water and reduced products
- Ferritin — sequesters free iron that would otherwise catalyze Fenton chemistry
This is the critical difference between Nrf2 activation and direct antioxidant supplementation: rather than supplying a fixed quantity of antioxidant molecules that get consumed in single-use reactions, Nrf2 activation raises the body's endogenous antioxidant production capacity. The increased glutathione synthesis, in particular, provides a continuously replenished pool of the body's primary intracellular antioxidant.
The Nrf2 pathway is the same pathway activated by sulforaphane (from broccoli sprouts), by exercise, by intermittent fasting, and by several other interventions associated with healthy aging. The convergence of these interventions on Nrf2 activation is part of why exercise, fasting, and polyphenol-rich diets all produce similar broad-spectrum health benefits.
For more on direct antioxidant interventions and the broader oxidative-stress literature, see our Oxidative Stress page.
COX-2 and iNOS Downregulation
Two NF-κB target genes deserve specific mention because they encode key enzymes in the inflammatory cascade: cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS). Both are strongly upregulated by NF-κB in response to inflammatory stimuli, and both contribute substantially to the tissue damage of chronic inflammation.
COX-2 produces prostaglandin E2 (PGE2), which mediates many of the cardinal signs of acute inflammation (heat, redness, swelling, pain) and contributes to chronic inflammation in arthritis, inflammatory bowel disease, and some cancers. The standard NSAIDs (ibuprofen, naproxen, diclofenac) inhibit both COX-1 (constitutive, gastric mucosal protective) and COX-2 (inducible, inflammatory). The selective COX-2 inhibitors (celecoxib, etoricoxib) target only COX-2 and avoid the gastric side effects of COX-1 inhibition. Resveratrol reduces COX-2 expression at the transcriptional level (via NF-κB suppression) and at the post-translational level (via direct enzyme inhibition at higher concentrations). The net effect is a selective COX-2 reduction without the COX-1 effects of traditional NSAIDs.
iNOS produces large quantities of nitric oxide in inflammatory cells (macrophages, neutrophils), where the NO combines with superoxide to form peroxynitrite, a potent oxidant that damages proteins, lipids, and DNA. Resveratrol downregulates iNOS expression at the transcriptional level (NF-κB suppression) and shifts the cellular NO production back toward the constitutive eNOS isoform that produces signaling-level NO in vascular endothelium (see our Cardiovascular Health deep-dive).
The selective effect on inducible (NF-κB-driven) inflammatory enzymes while sparing or even enhancing constitutive (homeostatic) enzymes is one of the favorable features of resveratrol's anti-inflammatory profile compared to broad-spectrum corticosteroids or non-selective NSAIDs.
Emodin (Anthraquinone) Effects
Emodin (1,3,8-trihydroxy-6-methylanthraquinone) is the principal anthraquinone in Japanese knotweed root, and it contributes a complementary set of effects to the herb's overall profile. Anthraquinones are a class of plant compounds with a three-ring quinone core, structurally related to the medicinal anthraquinones in rhubarb (Rheum officinale, knotweed's botanical cousin), senna (Senna alexandrina), cascara (Frangula purshiana), and aloe.
Emodin's documented pharmacological effects include:
- Anti-inflammatory — emodin inhibits NF-κB and downstream cytokine production through partly overlapping and partly distinct mechanisms from resveratrol. Combined, the two molecules produce stronger anti-inflammatory effects than either alone.
- Antifungal — in-vitro activity against Candida albicans and several other yeasts and filamentous fungi. The mechanism involves disruption of fungal cell membrane integrity and inhibition of mitochondrial function.
- Antibacterial — activity against Gram-positive bacteria (Staphylococcus aureus, Enterococcus faecalis), some Gram-negatives, and several anaerobes.
- Antiviral — in-vitro activity against several DNA and RNA viruses; the molecular mechanisms vary.
- Anti-tumor — preclinical activity against several cancer cell lines through apoptosis induction, cell cycle arrest, and reduced angiogenesis.
- Mild laxative — like other anthraquinones, emodin stimulates colonic motility through irritation of the colonic mucosa and enhanced peristalsis. This effect is the reason high doses of whole-root knotweed cause loose stools, and is the same mechanism behind the laxative effect of senna and cascara.
- Weak phytoestrogen-like activity — binds to estrogen receptors with low affinity, producing partial agonist/antagonist effects depending on tissue and baseline estrogen levels.
The phytoestrogen activity is weak enough not to be a primary clinical concern in most patients but strong enough to warrant caution in estrogen-sensitive cancers. The mild laxative effect is the principal reason that whole-root knotweed dosing has an upper ceiling determined by GI tolerance rather than pharmacological efficacy.
Traditional Chinese Medicine: "Blood Stasis" and Inflammation
Long before modern molecular biology, Traditional Chinese Medicine classified Japanese knotweed (hu zhang) as a herb for several distinct patterns, including:
- Damp-heat in the liver and gallbladder — manifesting as jaundice, hepatitis, gallstones, or related digestive disorders
- Blood stasis — the TCM term for the combination of impaired circulation, oxidation, and inflammation that manifests as bruising, traumatic injury sequelae, menstrual pain, and the typical pattern of chronic inflammatory disease
- Toxic heat — manifesting as snakebite reactions, burns, certain infections, and skin inflammation
- Lung-phlegm patterns — including persistent productive cough and bronchitis-like conditions
The TCM concept of "blood stasis" deserves particular attention because it maps so well onto the modern understanding of chronic inflammation. In TCM, blood stasis is a pattern characterized by fixed (rather than moving) pain, dark or purple tissue discoloration, dark-clotted menstrual blood, easy bruising, palpable masses, and a tendency to chronic disease that resists treatment. The classical herbs for blood stasis — including hu zhang, salvia (dan shen), persica (tao ren), carthamus (hong hua), and notoginseng (san qi) — share a common molecular profile when examined with modern pharmacology: they are anti-inflammatory, antioxidant, antiplatelet, vasodilatory, and microcirculation-supporting.
This convergence of traditional and modern understanding is one of the more interesting findings of contemporary ethnopharmacology. The Chinese physicians who classified hu zhang as a blood-stasis herb 1000+ years ago were observing the clinical effects we now describe in terms of NF-κB inhibition, antioxidant cascade activation, endothelial NO upregulation, and platelet inhibition. The framework was different; the underlying biology was the same.
For Western patients and clinicians, this means that the modern indications for Japanese knotweed (cardiovascular, anti-inflammatory, Lyme adjunct, longevity) overlap substantially with the traditional Chinese indications (blood stasis, chronic inflammation, hepatic disorders, post-traumatic recovery). The herb has been doing approximately the same set of things in approximately the same patient populations for many centuries, even as the conceptual framework has evolved.
Arthritis and Joint Inflammation
Joint inflammation is one of the most well-studied applications of resveratrol-rich preparations. The pathology of both osteoarthritis (OA) and rheumatoid arthritis (RA) involves chronic NF-κB-driven inflammation in the synovium, with downstream COX-2 upregulation, prostaglandin E2 production, MMP-driven cartilage degradation, and progressive joint damage.
Resveratrol acts on this pathology through:
- NF-κB inhibition in synoviocytes and infiltrating immune cells, reducing inflammatory cytokine production
- COX-2 downregulation and reduced prostaglandin E2 production (the NSAID mechanism, but selective)
- Reduced MMP-1, MMP-3, MMP-9, and MMP-13 expression, slowing cartilage degradation
- Direct chondroprotective effects on cartilage matrix synthesis
- Reduced osteoclast activation in adjacent bone, reducing periarticular bone loss
Clinical trials in OA have shown modest but consistent reductions in joint pain scores, improvements in WOMAC (Western Ontario and McMaster Universities Arthritis Index) function scores, and reduced need for rescue NSAID medication. The effect sizes are smaller than NSAIDs but with substantially better safety profiles for long-term use. Several small trials in RA have shown reductions in tender joint count, swollen joint count, and ESR/CRP markers when resveratrol is added to standard disease-modifying therapy.
The traditional Japanese name for knotweed — itadori, "remove pain" — reflects centuries of empirical observation that the herb reduces musculoskeletal pain. The modern mechanistic explanation (NF-κB inhibition, COX-2 reduction, MMP inhibition) is consistent with the traditional indication.
Inflammatory Bowel Disease
Inflammatory bowel disease (Crohn's disease and ulcerative colitis) is another condition where the NF-κB-driven mucosal inflammation provides a clear mechanistic match for resveratrol's anti-inflammatory profile. Several preclinical and clinical studies have examined resveratrol or knotweed extracts in IBD models:
- Dextran sulfate sodium (DSS) colitis in mice — resveratrol pretreatment reduces colitis severity, mucosal inflammation, cytokine production, and weight loss; the effect is dose-dependent
- TNBS colitis in rats — resveratrol reduces colonic damage and inflammatory cytokines
- Pilot clinical trials in ulcerative colitis — small trials of 500 mg resveratrol daily have shown reductions in NF-κB activation in colonic biopsies, reduced TNF-α production, and modest improvements in clinical activity scores
- Gut microbiome effects — resveratrol modulates the gut microbiome composition, increasing beneficial taxa (Akkermansia muciniphila, Lactobacillus, Bifidobacterium) and reducing inflammatory taxa
The clinical evidence is preliminary but mechanistically supported. Resveratrol is not a substitute for established IBD therapies (5-ASA agents, immunomodulators, biologics) but may be a reasonable adjunct, particularly for patients in remission who want to layer additional support onto their maintenance therapy.
One nuance for IBD patients: the anthraquinone content of whole-root knotweed can produce loose stools that might be confused with disease flare. Patients with IBD who use knotweed should consider purified resveratrol or polydatin extracts (which contain minimal anthraquinone) rather than whole-root preparations to avoid this confound.
Skin Inflammation and Atopic Dermatitis
Resveratrol's anti-inflammatory and antioxidant effects also apply to skin inflammation. Topical resveratrol preparations have been studied in atopic dermatitis, psoriasis-like dermatitis, and the inflammatory component of acne. Oral knotweed extract has been examined for systemic anti-inflammatory effects in cutaneous conditions.
Documented mechanisms in skin:
- NF-κB inhibition in keratinocytes and skin-resident immune cells (Langerhans cells, dermal dendritic cells)
- Reduced production of inflammatory cytokines and chemokines (TNF-α, IL-1β, IL-6, IL-17, CCL5, CXCL8)
- Modulated Th17/Treg balance in psoriasis-relevant models (analogous to the gut Th17/Treg effects discussed in our Vitamin A page)
- Reduced UV-induced skin inflammation and oxidative damage (relevant to photoaging)
- Improved skin barrier function through enhanced ceramide and filaggrin production
Topical resveratrol preparations are commercially available in cosmetic formulations marketed for anti-aging and antioxidant effects on photodamaged skin. The evidence for clinical-grade skin improvement is modest but real; the effect sizes are comparable to topical vitamin C and other antioxidant cosmetic ingredients.
For oral knotweed extract used systemically in cutaneous conditions, the effect sizes are smaller but applicable to a broader range of conditions because the systemic anti-inflammatory mechanism is not skin-specific.
Neuroinflammation and Microglial Modulation
Chronic neuroinflammation — primarily driven by microglial activation in response to age, infection, ischemia, traumatic injury, or accumulated misfolded proteins — is increasingly recognized as a central mechanism in Alzheimer's disease, Parkinson's disease, traumatic brain injury sequelae, and several chronic pain syndromes. Microglia are the brain's resident immune cells; their chronic activation produces sustained TNF-α, IL-1β, IL-6, and reactive oxygen species release into the brain parenchyma, with downstream neuronal damage.
Resveratrol crosses the blood-brain barrier (as detailed in our Lyme Disease deep-dive) and modulates microglial activation through:
- NF-κB inhibition in microglia, reducing pro-inflammatory cytokine production
- Shifting microglial polarization from M1 (pro-inflammatory) to M2 (tissue-repair) phenotype
- Reducing reactive oxygen species production by activated microglia
- Supporting astrocyte function and the blood-brain barrier integrity
- Direct neuroprotective effects on cortical and hippocampal neurons
Preclinical studies in Alzheimer's disease models have shown reduced amyloid-beta plaque burden, reduced neuroinflammation, and improved cognitive performance with chronic resveratrol administration. Small clinical trials in mild cognitive impairment and early Alzheimer's disease have shown reduced CSF inflammatory markers and stabilization of cognitive decline. The trials are too small and follow-up too short to establish clinical efficacy, but the mechanism and preliminary data justify ongoing study.
For more on the broader neuroinflammation framework and Alzheimer's disease specifically, see our Alzheimer's Disease page.
Oxidative Stress in Chronic Disease
Oxidative stress — the imbalance between reactive oxygen species production and antioxidant defenses — is a unifying mechanism in most chronic diseases of aging. The list of conditions in which oxidative stress is a documented driver of pathology is long: atherosclerosis, type 2 diabetes, neurodegenerative diseases, chronic kidney disease, COPD, fatty liver disease, rheumatologic disease, many cancers, and most autoimmune disease.
Resveratrol contributes to oxidative stress reduction through three mechanisms:
- Direct free radical scavenging — resveratrol's phenol groups donate hydrogen atoms to neutralize reactive species. The effect is real but limited — the molar concentration of resveratrol in tissue is too low for this to be the dominant mechanism.
- Nrf2 pathway activation — raises endogenous antioxidant production, providing a continuously replenished defense system. This is the most important mechanism for systemic effect.
- Mitochondrial protection — resveratrol supports mitochondrial biogenesis (via PGC-1α) and reduces mitochondrial ROS production by improving electron transport chain efficiency.
The clinical-trial evidence for resveratrol's effect on oxidative stress markers (MDA, F2-isoprostanes, oxidized LDL, urinary 8-OHdG) is consistent: modest reductions of 15 to 30% in elevated baselines, with smaller effects in already-low baselines. The reductions correlate with clinical improvements in the disease conditions where oxidative stress is mechanistically central.
For practical use, resveratrol from Japanese knotweed should be understood as a Nrf2 pathway activator rather than as a direct antioxidant. This mechanistic framing matches what is observed in clinical trials and avoids the (now-discredited) framing of antioxidant supplementation as a direct molar replacement for tissue antioxidant deficits.
For more on the broader oxidative stress framework and related interventions, see our Oxidative Stress page.
Cautions
- Anticoagulant/antiplatelet interactions — as discussed in the Cardiovascular Health deep-dive, resveratrol has measurable antiplatelet activity and compounds warfarin, DOACs, aspirin, clopidogrel, and other antiplatelet drugs. Stop 1 to 2 weeks before surgery.
- CYP3A4 inhibition — theoretical interaction with statins, calcium channel blockers, immunosuppressants, and many psychiatric medications. The clinical magnitude is uncertain but worth flagging to prescribers.
- Estrogen-sensitive cancers — both resveratrol and emodin have weak phytoestrogen activity. Use only under oncology supervision in active or recent estrogen receptor-positive breast or endometrial cancer.
- Pregnancy and lactation — insufficient safety data; avoid in the absence of specific clinical indication.
- Anthraquinone-related GI effects — whole-root knotweed and lower-percentage standardized extracts retain emodin and other anthraquinones, which can cause loose stools or diarrhea at high doses. Reduce dose if this develops. Purified resveratrol products avoid this issue.
- Chronic high-dose hepatotoxicity — rare case reports of liver injury at gram-range chronic doses. Baseline liver enzymes are reasonable before long-term high-dose use.
- Drug-induced laxative dependence — chronic use of anthraquinone-containing products (knotweed, senna, cascara) can produce melanosis coli and laxative dependence. Use whole-root knotweed in short courses or pulse-dose; chronic continuous high-dose use should favor purified extracts.
- Oxalate content — whole-root knotweed is high in oxalate. Patients with calcium-oxalate kidney stones should prefer purified resveratrol or polydatin extracts.
- Iron absorption — polyphenols including resveratrol can reduce non-heme iron absorption from food. Separate by 2 hours from iron-rich meals or iron supplements.
Key Research Papers
- Manna SK, Mukhopadhyay A, Aggarwal BB (2000). Resveratrol suppresses TNF-induced activation of nuclear transcription factors NF-κB, activator protein-1, and apoptosis. J Immunol 164:6509-6519. — PubMed
- Yeung F, Hoberg JE, Ramsey CS et al. (2004). Modulation of NF-κB-dependent transcription and cell survival by the SIRT1 deacetylase. EMBO J 23:2369-2380. — PubMed
- Chen LF, Mu Y, Greene WC (2002). Acetylation of RelA at discrete sites regulates distinct nuclear functions of NF-κB. EMBO J. — PubMed
- Kode A, Rajendrasozhan S, Caito S et al. (2008). Resveratrol induces glutathione synthesis by activation of Nrf2 and protects against cigarette smoke-mediated oxidative stress in human lung epithelial cells. Am J Physiol Lung Cell Mol Physiol. — PubMed
- Subbaramaiah K, Chung WJ, Michaluart P et al. (1998). Resveratrol inhibits cyclooxygenase-2 transcription and activity in phorbol ester-treated human mammary epithelial cells. J Biol Chem. — PubMed
- Tsai SH, Lin-Shiau SY, Lin JK (1999). Suppression of nitric oxide synthase and the down-regulation of the activation of NF-κB in macrophages by resveratrol. Br J Pharmacol. — PubMed
- Dong Z (2003). Molecular mechanism of the chemopreventive effect of resveratrol. Mutat Res 523-524:145-150. — PubMed
- Dolinsky VW, Dyck JR (2011). Calorie restriction and resveratrol in cardiovascular health and disease. Biochim Biophys Acta. — PubMed
- Singh CK, Liu X, Ahmad N (2015). Resveratrol, in its natural combination in whole grape, for health promotion and disease management. Ann N Y Acad Sci. — PubMed
- Movahed A, Raj P, Nabipour I et al. (2020). Efficacy and safety of resveratrol in type 1 diabetes patients. Nutrients. — PubMed
- Srivastava AK et al. Emodin: a review of its pharmacology, toxicity, and pharmacokinetics. Phytother Res. — PubMed
- Surh YJ (2003). Cancer chemoprevention with dietary phytochemicals. Nat Rev Cancer. — PubMed
PubMed Topic Searches
- PubMed: Resveratrol and NF-κB inhibition
- PubMed: SIRT1 deacetylation of p65
- PubMed: Resveratrol and Nrf2
- PubMed: Resveratrol and arthritis
- PubMed: Resveratrol and IBD
- PubMed: Emodin pharmacology
- PubMed: Polygonum cuspidatum anti-inflammatory
Connections
- Japanese Knotweed Overview
- Japanese Knotweed Benefits Hub
- Knotweed Resveratrol & Longevity
- Knotweed for Lyme Disease
- Knotweed for Cardiovascular Health
- Oxidative Stress
- Anti-Inflammatory Diet
- Longevity Protocols
- Turmeric (Curcumin)
- Berberine
- Astragalus
- Cat's Claw
- Andrographis
- Atherosclerosis
- Crohn's Disease
- Ulcerative Colitis
- Alzheimer's Disease
- Metabolic Syndrome
- Rheumatoid Arthritis
- All Herbs