Gut-Brain Axis and Vagal Nerve Signaling
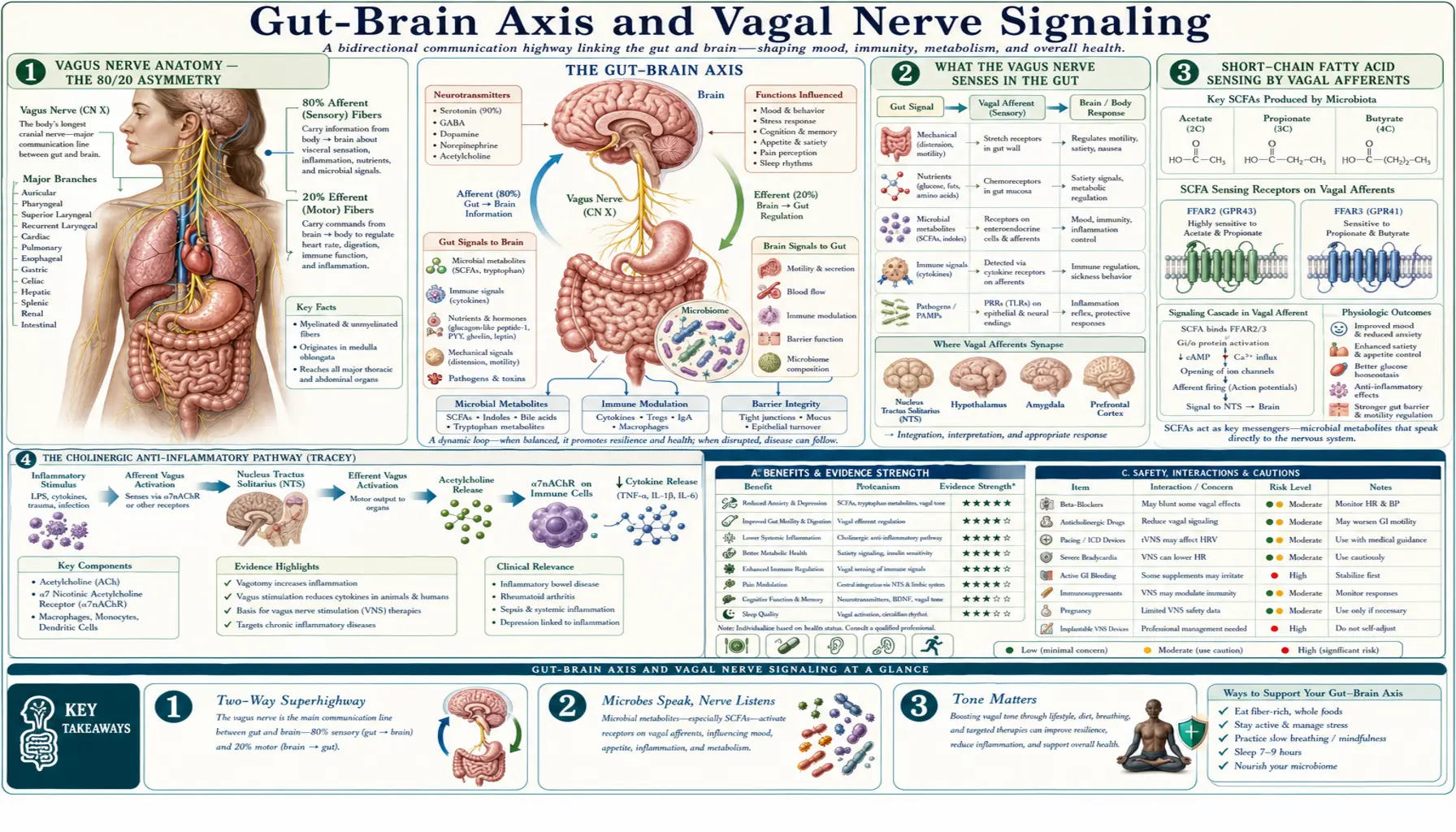
The vagus nerve (cranial nerve X) is the longest cranial nerve in the body and the dominant parasympathetic conduit between the gut and the brain. Roughly 80% of vagal fibers are afferent (carrying information from gut to brain), and only 20% are efferent (carrying signals from brain to gut). This asymmetry is the key anatomical fact behind the gut-brain axis: your gut is talking to your brain far more than your brain is talking to your gut. Vagal afferents detect microbial metabolites, mechanical distension, gut peptides, and inflammatory signals, integrating them at the nucleus tractus solitarius and projecting onward to the locus coeruleus, raphe nuclei, amygdala, and prefrontal cortex — brain regions that govern mood, arousal, and stress reactivity. This page walks through the anatomy, signaling mechanisms, the FDA-cleared therapeutic applications of vagus nerve stimulation, and what current evidence supports about non-invasive vagal tone training.
Table of Contents
- Vagus Nerve Anatomy — The 80/20 Asymmetry
- What the Vagus Nerve Senses in the Gut
- Short-Chain Fatty Acid Sensing by Vagal Afferents
- The Cholinergic Anti-Inflammatory Pathway (Tracey)
- Vagotomy Experiments — Proving the Vagus is the Conduit
- FDA-Cleared Vagus Nerve Stimulation for Depression
- Transcutaneous Vagus Nerve Stimulation (tVNS)
- Heart Rate Variability as Biomarker of Vagal Tone
- Non-Invasive Vagal Tone Training (Practical)
- The Vagus Nerve and Parkinson's Disease
- Key Research Papers
- Connections
- Featured Videos
Vagus Nerve Anatomy — The 80/20 Asymmetry
The vagus nerve emerges from the medulla oblongata, exits the skull through the jugular foramen, and descends through the neck alongside the carotid artery, branching to innervate the larynx, pharynx, heart, lungs, esophagus, stomach, pancreas, liver, gallbladder, small intestine, and proximal colon (roughly through the splenic flexure; the distal colon and rectum are innervated by sacral parasympathetics). The name "vagus" comes from the Latin word for "wandering" — the nerve's territory is unusually extensive for a single cranial nerve.
The most important anatomical fact about the vagus for gut-brain signaling is that it is approximately 80% afferent — the fibers carry sensory information from the periphery toward the brainstem — and only 20% efferent. Most clinical and popular discussion of the vagus emphasizes its efferent parasympathetic role ("rest and digest"), but the anatomical reality is that the vagus is primarily a sensor, not an effector. Information flow is overwhelmingly bottom-up: the gut is informing the brain.
Vagal afferent cell bodies sit in the nodose ganglion, just outside the skull. Their central terminals project to the nucleus tractus solitarius (NTS) in the dorsal medulla, which is the first relay station. From NTS, projections ascend to the parabrachial nucleus, the hypothalamic paraventricular nucleus (PVN, the apex of the HPA stress axis), the locus coeruleus (the noradrenergic arousal center), the dorsal raphe (the serotonergic mood center), the amygdala (fear and emotional salience), and the insular and prefrontal cortices (interoception and executive function). This explains anatomically why a gut-derived signal can modulate mood, arousal, stress, and cognition — the wire reaches all of them.
What the Vagus Nerve Senses in the Gut
Vagal afferent terminals in the gut wall do not contact the lumen directly. They sit in the lamina propria and the muscle layers, sensing through several intermediary mechanisms:
- Mechanoreceptors — intraganglionic laminar endings and intramuscular arrays detect distension and contraction of the gut wall. These provide the satiety signal that follows a meal and contribute to gastric stretch reception.
- Chemoreceptors via enteroendocrine cells — specialized epithelial cells (enterochromaffin cells, L cells, I cells, K cells) sense luminal nutrients, microbial metabolites, and bile acids, then release signaling molecules (serotonin, GLP-1, CCK, GIP, ghrelin) that activate vagal afferent terminals in the underlying lamina propria. Many enteroendocrine cells form direct synapse-like contacts ("neuropods") with vagal terminals, allowing sub-second signaling, not just paracrine diffusion.
- Immune sensing — cytokines released by mucosal immune cells (especially IL-6, IL-1-beta, TNF-alpha) activate vagal afferents during inflammation. This is the afferent arm of the cholinergic anti-inflammatory reflex.
- Bacterial product sensing — vagal afferents respond to bacterial lipopolysaccharide and to short-chain fatty acids produced by colonic fermentation, either directly or via enterocyte-released mediators.
The integrated effect is that the vagus is providing the brain with a continuous status report on luminal nutrient content, gut wall tension, mucosal immune state, and microbial activity. Most of this information never reaches conscious awareness — it operates entirely within the autonomic and interoceptive nervous system.
Short-Chain Fatty Acid Sensing by Vagal Afferents
Short-chain fatty acids (SCFAs) — primarily butyrate, propionate, and acetate — are produced when colonic bacteria ferment dietary fiber. They are the dominant energy source of colonocytes (butyrate provides up to 70% of colonocyte energy), but they are also key signaling molecules in the gut-brain axis. SCFAs bind to free fatty acid receptors FFAR2 (GPR43) and FFAR3 (GPR41) expressed on enteroendocrine cells and on vagal afferent terminals themselves.
SCFA-driven vagal signaling explains several non-obvious connections. Diets high in fermentable fiber produce more butyrate, more PYY and GLP-1 release, stronger vagal afferent firing, and stronger satiety signaling at the hypothalamic PVN. Disrupting butyrate-producing bacteria (the Faecalibacterium prausnitzii and Roseburia genera are the dominant producers in healthy humans) by antibiotic exposure or Western dietary patterns reduces this signaling stream. In murine models, depleting Faecalibacterium reduces vagal afferent activity and produces depressive-like behavior; restoring butyrate-producing bacteria rescues both.
For practical clinical relevance, the takeaway is that dietary fiber is not just bulk — it is the substrate for a bacterial signaling molecule that the vagus nerve actively monitors. This is one of the strongest mechanistic arguments for the Mediterranean-style high-fiber diet as a depression and anxiety intervention, beyond the more familiar inflammation arguments.
The Cholinergic Anti-Inflammatory Pathway (Tracey)
Kevin Tracey's laboratory at the Feinstein Institute identified, in a landmark 2002 Nature paper, that vagal efferent stimulation suppresses systemic inflammation through release of acetylcholine onto macrophages bearing the alpha-7 nicotinic acetylcholine receptor (alpha7-nAChR). Acetylcholine binding to alpha7-nAChR inhibits NF-kB activation in the macrophage and dramatically reduces release of TNF-alpha, IL-1-beta, and IL-6.
This is the "cholinergic anti-inflammatory pathway." Its anatomy is largely splenic — vagal efferent fibers do not directly innervate the spleen, but they activate splenic norepinephrine-releasing fibers that in turn activate T cells expressing choline acetyltransferase, which release acetylcholine onto splenic macrophages. The pathway constitutes a real-time, neural mechanism for the brain to dampen systemic inflammation.
Therapeutically, this pathway is now being exploited by implantable vagus nerve stimulators in rheumatoid arthritis (SetPoint Medical trials), Crohn's disease, and Lupus. Pilot trials show meaningful clinical improvement in patients refractory to standard biologic therapy. The mechanism also explains why low vagal tone (low heart rate variability) is consistently associated with elevated systemic inflammatory markers (CRP, IL-6) in observational studies, and why vagal-tone-improving interventions reduce inflammation.
Vagotomy Experiments — Proving the Vagus is the Conduit
The most rigorous evidence that the vagus is the actual physical conduit for gut-to-brain signaling comes from vagotomy experiments. The classic Bravo, Forsythe, et al. (2011) PNAS paper at McMaster University fed mice Lactobacillus rhamnosus JB-1 and showed reduced anxiety-like behavior, reduced corticosterone response to stress, and altered GABA receptor expression in the brain. The critical control: surgical vagotomy completely abolished the effect. The probiotic did not work without an intact vagus nerve.
This was the first conclusive demonstration that a specific microbial intervention produces a CNS effect through a defined neural pathway, not through a diffusible blood-borne mediator. Subsequent work has shown that vagotomy abolishes most probiotic effects on mood and behavior, but does not abolish all microbiome-driven CNS effects — SCFA effects on microglial development, for instance, appear to operate partly via direct circulation. The vagus is one of several pathways but is clearly the dominant rapid-signaling channel.
Human evidence is necessarily more indirect (we cannot randomize humans to vagotomy), but a striking 2015 Danish epidemiologic study (Svensson et al., Annals of Neurology) found that patients who had undergone full-truncal vagotomy for peptic ulcer disease in the 1970s and 1980s had significantly reduced lifetime risk of Parkinson's disease (HR approximately 0.6) compared to matched controls with partial vagotomy or no vagotomy. This is consistent with the Braak hypothesis that Parkinson's alpha-synuclein pathology originates in the enteric nervous system and ascends to the brainstem along vagal fibers.
FDA-Cleared Vagus Nerve Stimulation for Depression
Implantable vagus nerve stimulation (VNS) was originally FDA-approved in 1997 for refractory epilepsy. The neurosurgical procedure implants a pacemaker-like device in the left chest wall with a lead that wraps around the left cervical vagus nerve. The device delivers programmable electrical pulses (typically 0.25-3.0 mA, 250-500 microsecond pulse width, 30 seconds on, 5 minutes off).
In 2005, VNS received FDA approval for adjunctive treatment of chronic or recurrent treatment-resistant major depressive disorder in adults with at least four failed antidepressant trials. Response rates are modest in the first year (approximately 15-20% response rate at 12 weeks) but rise substantially with continued stimulation — observational follow-up of patients implanted for depression shows response rates of 40-50% at 12-24 months, with sustained benefit for years. Medicare coverage was finally extended in 2019 after a long battle over evidence interpretation.
The mechanism of VNS antidepressant action is thought to involve upregulation of norepinephrine in the locus coeruleus, serotonin in the dorsal raphe, increased BDNF expression, normalization of HPA axis hyperactivity, and reduced systemic inflammation via the cholinergic anti-inflammatory pathway. The clinical signature is a slow onset (weeks to months) but durable response — quite different from rapid-acting antidepressants like ketamine.
Transcutaneous Vagus Nerve Stimulation (tVNS)
The auricular branch of the vagus nerve (the only cutaneous representation of the vagus) innervates a small territory of the external ear — specifically the concha and the tragus. This anatomy enables non-invasive transcutaneous vagus nerve stimulation (tVNS) using surface electrodes clipped to the ear, avoiding the need for surgical implantation.
tVNS has been studied in:
- Major depressive disorder — multiple small RCTs show modest antidepressant effects (effect sizes 0.3-0.5), with the gammaCore handheld device receiving FDA clearance for migraine and cluster headache. Larger depression trials are ongoing.
- Migraine — FDA-cleared for acute treatment of migraine and cluster headache.
- Treatment-resistant epilepsy — non-invasive alternative to surgical VNS.
- Tinnitus — emerging evidence for combined tVNS plus auditory rehabilitation.
- Long COVID, POTS, and dysautonomia — preliminary trials of tVNS for autonomic symptoms.
- Inflammatory bowel disease and rheumatoid arthritis — small open-label studies exploring the cholinergic anti-inflammatory pathway.
Consumer-grade tVNS devices are now widely available without prescription (Nurosym, Pulsetto, Xen by Neuvana). The evidence base for consumer devices is much thinner than for clinical-grade devices used in trials, and stimulation parameters vary widely. For patients interested in tVNS, the safest path is participation in a clinical trial or use of an FDA-cleared device for an approved indication.
Heart Rate Variability as Biomarker of Vagal Tone
Heart rate variability (HRV) is the variation in time between consecutive heartbeats. The high-frequency component of HRV (HF-HRV, 0.15-0.4 Hz) reflects respiratory sinus arrhythmia — the natural acceleration of heart rate during inhalation and deceleration during exhalation — which is mediated almost entirely by vagal efferent activity to the sinoatrial node. HF-HRV is therefore one of the few non-invasive measures of true vagal tone available.
Higher HRV is robustly associated with:
- Lower all-cause mortality and cardiovascular mortality
- Lower systemic inflammatory markers (CRP, IL-6)
- Better cognitive flexibility and executive function
- Lower rates of major depression and anxiety
- Better emotion regulation and stress recovery
- Higher gut microbiome diversity
HRV can be measured by chest-strap heart rate monitors (Polar H10), photoplethysmography wrist devices (Oura, WHOOP, Apple Watch), or finger pulse oximeters. The most reliable metric for vagal tone is the RMSSD (root mean square of successive differences). Healthy adults typically have RMSSD in the 30-70 millisecond range; below 20 ms suggests significantly reduced vagal tone. Daily morning measurement is more useful than spot measurement because HRV varies substantially with sleep, stress, and recent physical exertion.
Non-Invasive Vagal Tone Training (Practical)
Several non-invasive interventions have been shown to acutely or chronically increase vagal tone as measured by HRV. The evidence base ranges from solid (slow breathing, exercise) to suggestive (cold exposure, gargling, humming):
- Slow paced breathing at ~6 breaths per minute — this matches the natural Mayer wave frequency and produces maximal respiratory sinus arrhythmia. Roughly 5 seconds inhale, 5 seconds exhale. Practiced 10-20 minutes daily, this is the single best-validated vagal tone training intervention. Apps: Breathwrk, Calm, Othership.
- Cold exposure — cold water face immersion (the diving reflex), cold showers, and ice baths acutely shift the autonomic balance toward parasympathetic dominance. The Wim Hof method combines breathwork and cold exposure. Evidence for chronic adaptation is weaker than evidence for acute effect.
- Aerobic exercise — sustained aerobic training increases resting HRV and reduces resting heart rate. The relationship is dose-dependent and quite reproducible across trials.
- Yoga, tai chi, and qigong — meta-analyses show modest HRV increases with regular practice, likely mediated by the combined effects of slow breathing, mild physical activity, and stress reduction.
- Meditation, especially loving-kindness and compassion meditation — multiple trials show HRV increases during and immediately after practice, with smaller but persistent effects with daily practice.
- Singing, humming, and gargling — activate the laryngeal and pharyngeal branches of the vagus. Evidence is thin but mechanistically plausible.
- Sleep optimization — sleep deprivation acutely lowers HRV; consistent sleep schedules with adequate duration restore it.
- Reducing chronic stress, dietary inflammation, and alcohol — all three depress HRV; reducing them tends to restore it.
For patients with documented low vagal tone and gut-brain symptoms (IBS with anxiety, depression with constipation, chronic fatigue with dysautonomia), a daily 10-minute slow-breathing practice combined with regular aerobic exercise is a reasonable first-line intervention with no cost and no downside. See our Restoration Protocols page for integration with dietary and probiotic interventions.
The Vagus Nerve and Parkinson's Disease
The Braak hypothesis (Heiko Braak, 2003) proposes that Parkinson's disease pathology originates not in the substantia nigra but in the enteric nervous system and the olfactory bulb, then ascends along the vagus nerve to the dorsal motor nucleus of the vagus in the medulla, then progresses through the brainstem to the substantia nigra and onward to the cortex. Alpha-synuclein aggregates can be found in the enteric nervous system years before motor symptoms appear, and constipation is one of the earliest premotor symptoms of Parkinson's, often preceding the motor diagnosis by a decade or more.
The Svensson 2015 vagotomy epidemiology (cited above) and subsequent rodent work using alpha-synuclein pre-formed fibrils injected into the gut wall and shown to propagate to the brain via the vagus — with vagotomy preventing propagation — have largely confirmed the Braak hypothesis at the mechanistic level. The gut microbiome may be the upstream trigger; certain bacterial protein products (curli from E. coli, for instance) can cross-seed alpha-synuclein aggregation.
The clinical implications are still being worked out, but suggest that gut-targeted interventions in early-stage Parkinson's (or in high-risk pre-clinical individuals) may slow disease progression. For more on Parkinson's management see our Parkinson's Disease page.
Key Research Papers
- Bravo JA, Forsythe P, Chew MV, Escaravage E, Savignac HM, Dinan TG, Bienenstock J, Cryan JF (2011). Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. PNAS 108(38):16050-16055. — PubMed: Bravo 2011
- Borovikova LV, Ivanova S, Zhang M, Yang H, Botchkina GI, Watkins LR, Wang H, Abumrad N, Eaton JW, Tracey KJ (2000). Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 405:458-462. — Search PubMed
- Tracey KJ (2002). The inflammatory reflex. Nature 420:853-859. — PubMed: Tracey inflammatory reflex
- Svensson E, Horvath-Puho E, Thomsen RW, Djurhuus JC, Pedersen L, Borghammer P, Sorensen HT (2015). Vagotomy and subsequent risk of Parkinson's disease. Annals of Neurology 78(4):522-529. — Search PubMed
- Bonaz B, Bazin T, Pellissier S (2018). The vagus nerve at the interface of the microbiota-gut-brain axis. Frontiers in Neuroscience 12:49. — PubMed: Bonaz review
- Rush AJ, Marangell LB, Sackeim HA, et al. (2005). Vagus nerve stimulation for treatment-resistant depression: a randomized, controlled acute phase trial. Biological Psychiatry 58(5):347-354. — PubMed: Rush VNS depression
- Forsythe P, Bienenstock J, Kunze WA (2014). Vagal pathways for microbiome-brain-gut axis communication. Advances in Experimental Medicine and Biology 817:115-133. — PubMed: Forsythe vagal pathways
- Thayer JF, Lane RD (2009). Claude Bernard and the heart-brain connection: further elaboration of a model of neurovisceral integration. Neuroscience & Biobehavioral Reviews 33(2):81-88. — PubMed: Thayer HRV
- Liu B, Fang F, Pedersen NL, et al. (2017). Vagotomy and Parkinson disease: a Swedish register-based matched-cohort study. Neurology 88(21):1996-2002. — PubMed: Liu Swedish cohort
- Kaczmarczyk R, Tejera D, Simon BJ, Heneka MT (2017). Microglia modulation through external vagus nerve stimulation in a murine model of Alzheimer's disease. Journal of Neurochemistry. — Search PubMed
- Koopman FA, Chavan SS, Miljko S, et al. (2016). Vagus nerve stimulation inhibits cytokine production and attenuates disease severity in rheumatoid arthritis. PNAS 113(29):8284-8289. — PubMed: Koopman RA
- Yuan H, Silberstein SD (2016). Vagus nerve and vagus nerve stimulation, a comprehensive review. Headache 56(1):71-78. — Search PubMed
PubMed Topic Searches
- PubMed: VNS treatment-resistant depression
- PubMed: Transcutaneous VNS
- PubMed: HRV vagal tone inflammation
- PubMed: Cholinergic anti-inflammatory pathway
- PubMed: Vagus Parkinson's Braak
Connections
- Gut-Brain Axis Hub
- Gut-Brain Benefits Deep-Dive
- Serotonin and Mood
- Stress and Permeability
- Restoration Protocols
- Stress Management
- Natural Anxiety Relief
- Gut Healing
- Depression
- Anxiety
- Parkinson's Disease
- Migraine
- Irritable Bowel Syndrome
- Fermented Foods
- All Remedies