Gut-Brain Axis, Serotonin, and Mood
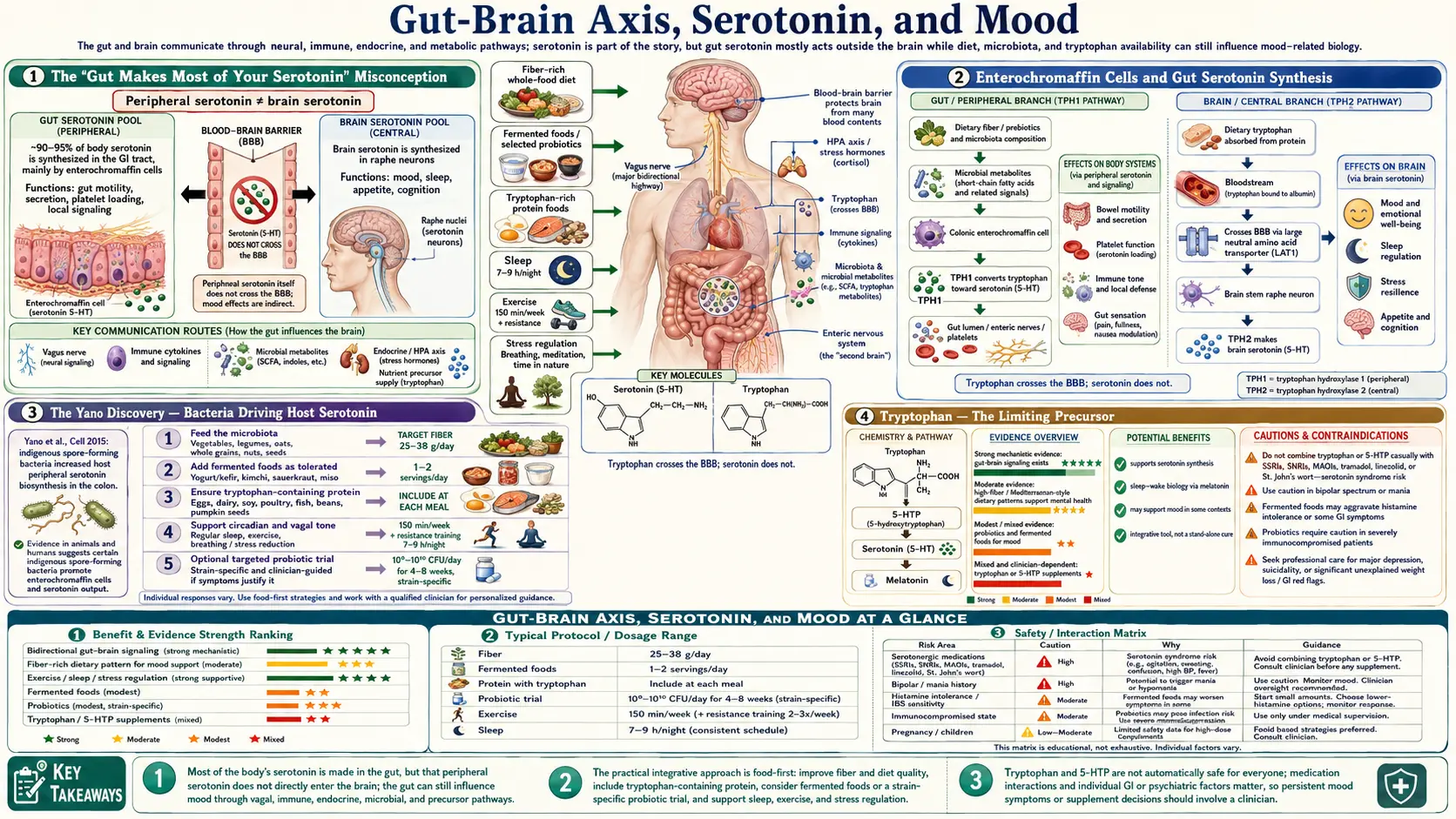
Approximately 90-95% of body serotonin is produced not in the brain but in enterochromaffin cells of the gastrointestinal epithelium. This is the central, frequently-cited fact of the gut-brain axis — and also the most frequently misunderstood. Gut-derived serotonin does not cross the blood-brain barrier and therefore cannot directly raise brain serotonin. What it does do is modulate gut motility, vagal afferent firing, mucosal immunity, and the production of brain-relevant tryptophan metabolites that can cross the blood-brain barrier and influence mood. The microbiome shapes this entire system — certain bacterial species drive enterochromaffin cell serotonin synthesis, others divert tryptophan into the kynurenine pathway under inflammation, and a third group produce GABA, dopamine precursors, and other neuroactive molecules directly. This page walks through the actual mechanisms, the strain-specific psychobiotic evidence, and what the gut serotonin story means — and does not mean — for depression and anxiety treatment.
Table of Contents
- The "Gut Makes Most of Your Serotonin" Misconception
- Enterochromaffin Cells and Gut Serotonin Synthesis
- The Yano Discovery — Bacteria Driving Host Serotonin
- Tryptophan — The Limiting Precursor
- The Kynurenine Pathway Diversion Under Inflammation
- Psychobiotics — Bacteria With Defined Mood Effects
- GABA and Dopamine-Precursor Producing Bacteria
- Why SSRIs Cause Gut Side Effects
- Fecal Microbiota Transplantation for Depression
- Clinical Implications for Mood Disorders
- Key Research Papers
- Connections
- Featured Videos
The "Gut Makes Most of Your Serotonin" Misconception
The statement that "90-95% of your serotonin is made in the gut" is true but routinely misinterpreted. It is often presented as "fix your gut and you fix your serotonin and you fix your depression." That logical chain skips two critical facts: (1) gut serotonin does not cross the blood-brain barrier, and (2) brain serotonin is synthesized independently in the dorsal raphe nucleus from circulating tryptophan that does cross the blood-brain barrier.
The gut and brain serotonin pools are effectively separate compartments. They use the same molecule and the same biosynthetic enzyme (tryptophan hydroxylase — TPH1 in the periphery, TPH2 in the brain — two different gene products with different regulation), but they do not directly exchange serotonin. Raising gut serotonin by activating enterochromaffin cells does not raise brain serotonin in the way that a brain-targeted SSRI does.
What gut serotonin does do is mechanistically important and indirectly influences brain function through several distinct pathways: it modulates gut motility via 5-HT3 and 5-HT4 receptors on enteric neurons, it activates vagal afferents that signal interoceptively to the brainstem and limbic system, it modulates mucosal immune responses, and most importantly, it influences the systemic availability of tryptophan — the rate-limiting precursor for brain serotonin. The gut-brain serotonin story is real, but the mechanism is not the simple plumbing one most popular accounts suggest.
Enterochromaffin Cells and Gut Serotonin Synthesis
Enterochromaffin (EC) cells are a subtype of enteroendocrine cell scattered throughout the gut epithelium, most densely in the duodenum and distal small intestine, with a smaller population in the colon. They were named in the late 19th century for their ability to be stained by chromium salts (chromaffin reaction), which we now know reflects their serotonin content.
EC cells synthesize serotonin from tryptophan using tryptophan hydroxylase 1 (TPH1) to produce 5-hydroxytryptophan (5-HTP), then aromatic L-amino acid decarboxylase to produce serotonin. The serotonin is packaged into secretory vesicles and released in response to luminal stimuli — nutrients (especially carbohydrates and fats), bile acids, microbial metabolites (especially short-chain fatty acids and indole derivatives), and mechanical stretch.
Released serotonin acts locally on:
- Enteric neurons — 5-HT3 receptors on intrinsic primary afferent neurons initiate peristalsis; 5-HT4 receptors enhance neurotransmitter release at enteric synapses (the basis for the 5-HT4 agonist drug class, prucalopride, for chronic constipation).
- Vagal afferent terminals — 5-HT3 receptors transduce gut-to-brain signaling about nutrient content; this is why 5-HT3 antagonists (ondansetron, the "-setron" class) are effective antiemetics — they block the gut-to-brainstem nausea signal.
- Mucosal immune cells — mast cells, T cells, and dendritic cells all express serotonin receptors and respond to gut serotonin.
- Platelets — circulating platelets actively take up gut-derived serotonin via the serotonin transporter (SERT) and store it; platelet serotonin is what is measured in standard blood serotonin tests, and it reflects gut production, not brain production.
For context: the entire human body contains about 10 milligrams of serotonin, of which roughly 9.5 milligrams is in the gut and platelet pool, and only about 0.5 milligrams is in the brain. The brain serotonin pool is small but functionally critical.
The Yano Discovery — Bacteria Driving Host Serotonin
One of the landmark gut-brain axis papers is Yano JM et al. (2015) in Cell, from the Mazmanian laboratory at Caltech. The team showed that germ-free mice (raised without any gut bacteria) have approximately 50% lower colonic serotonin levels than conventionally raised mice. Colonizing the germ-free mice with a defined community of spore-forming bacteria (primarily Clostridium species, which are part of the normal human gut flora) restored colonic serotonin to normal levels.
The mechanism: spore-forming bacteria produce specific metabolites (deoxycholate, butyrate, propionate, and others) that signal enterochromaffin cells to upregulate TPH1 expression and serotonin synthesis. This was the first conclusive evidence that the gut microbiome actively drives host serotonin production, not merely correlating with it.
Subsequent work has extended this to specific human-relevant bacteria. Akkermansia muciniphila (a mucin-degrading bacterium associated with metabolic health), several Bifidobacterium species, and certain Lactobacillus strains all enhance host serotonin synthesis. Conversely, antibiotic-induced dysbiosis acutely reduces colonic serotonin in mouse models, with measurable changes in gut motility and behavior.
The clinical translation is still preliminary but suggests that microbiome interventions may modulate gut serotonin signaling in functional GI disorders (IBS-C and IBS-D are characterized by altered gut serotonin signaling) and possibly in the autonomic and mood comorbidities of those disorders.
Tryptophan — The Limiting Precursor
Tryptophan is one of the nine essential amino acids — humans cannot synthesize it and must obtain it from dietary protein. It is also one of the least abundant amino acids in most protein sources (roughly 1% of total amino acid content), and the rate-limiting precursor for both gut and brain serotonin synthesis.
Of dietary tryptophan absorbed in the small intestine:
- The majority is incorporated into body protein synthesis (the dominant fate, accounting for >95% in adults)
- A small fraction is metabolized by enterochromaffin cells for gut serotonin synthesis (typically <1% of intake)
- A larger fraction is metabolized by the kynurenine pathway in the liver and immune cells (see next section)
- A small fraction crosses the blood-brain barrier via the large neutral amino acid transporter (LAT1), where it is the precursor for brain serotonin synthesis
The blood-brain barrier transport is competitive — tryptophan shares LAT1 with the other large neutral amino acids (phenylalanine, tyrosine, leucine, isoleucine, valine, methionine, histidine). A meal high in branched-chain amino acids (BCAAs, common in whey protein and animal protein generally) actually reduces brain tryptophan transport because the BCAAs compete for the transporter. This is why classic 1970s research found that carbohydrate-rich meals raise brain serotonin (carbohydrates trigger insulin, which drives BCAAs into muscle, lowering their plasma concentration and freeing the transporter for tryptophan) while pure-protein meals can paradoxically lower brain tryptophan.
This is also the mechanism behind the experimental "acute tryptophan depletion" technique used in psychiatric research — subjects drink an amino acid mixture containing all essential amino acids except tryptophan, which forces protein synthesis to use up the body's tryptophan stores within hours and produces a measurable drop in CSF 5-HIAA. The technique reliably induces low mood in recovered depressed patients and in healthy individuals with a family history of depression, confirming the causal role of brain serotonin in mood vulnerability. See our L-Tryptophan page for more on supplementation.
The Kynurenine Pathway Diversion Under Inflammation
The kynurenine pathway is the dominant non-protein-synthesis fate of dietary tryptophan, accounting for the majority of tryptophan catabolism. It is regulated primarily by two enzymes: tryptophan-2,3-dioxygenase (TDO) in the liver, and indoleamine-2,3-dioxygenase (IDO) in immune cells and other peripheral tissues.
The critical regulatory point: IDO is strongly induced by inflammatory cytokines, particularly interferon-gamma, TNF-alpha, and IL-6. In the context of chronic systemic inflammation (whether from infection, autoimmune disease, obesity, leaky gut, or chronic stress), IDO induction shunts a much larger fraction of dietary tryptophan into the kynurenine pathway. The downstream consequences:
- Less tryptophan available for brain serotonin synthesis — mood-lowering effect
- Accumulation of neuroactive kynurenine metabolites — specifically quinolinic acid (a potent NMDA receptor agonist, neurotoxic at high concentrations) and 3-hydroxykynurenine (oxidative stress)
- Some neuroprotective metabolites also — kynurenic acid is an NMDA receptor antagonist with potentially protective effects
This is the "kynurenine hypothesis of depression": chronic inflammation diverts tryptophan into kynurenine, producing both relative brain serotonin deficiency and elevated neurotoxic quinolinic acid. The hypothesis is supported by:
- Elevated peripheral quinolinic acid in major depression and especially in inflammation-associated depression
- Higher rates of depression in conditions with chronic inflammation (obesity, autoimmune disease, post-stroke, post-MI, HIV)
- Reduced kynurenine pathway activation and improved mood with anti-inflammatory interventions
- Improved depression scores in some trials of anti-inflammatory agents (omega-3 fatty acids, anti-TNF biologics in psoriasis)
The gut connection: gut barrier breakdown (leaky gut, discussed in detail on the Stress and Permeability page) allows bacterial lipopolysaccharide translocation, which drives systemic inflammation, which induces IDO, which diverts tryptophan, which lowers brain serotonin. This is one of the most fully worked-out mechanistic chains linking gut health to mood.
Psychobiotics — Bacteria With Defined Mood Effects
The term "psychobiotics" was coined by Ted Dinan and John Cryan at the APC Microbiome Ireland to refer to live microorganisms that, when ingested in adequate amounts, confer mental health benefits. The evidence base is now substantial enough to identify specific strains with documented effects:
- Lactobacillus rhamnosus JB-1 — the strain used in the Bravo 2011 PNAS vagotomy study. Reduces anxiety-like behavior and corticosterone response in rodents; effect abolished by vagotomy. Limited human data.
- Bifidobacterium longum 1714 — randomized controlled trials in healthy human volunteers (Allen 2016, Translational Psychiatry) showed reduced cortisol response to social stress test, reduced anxiety self-report, and improved memory after 4 weeks of supplementation.
- Lactobacillus helveticus R0052 plus Bifidobacterium longum R0175 — the formulation marketed as Probio'Stick or Cerebiome. Multiple human RCTs show modest reductions in anxiety and depression scores. Effect sizes are small but consistent.
- VSL#3 / Visbiome (8-strain mixture) — primarily studied for ulcerative colitis and pouchitis, with secondary effects on mood scores in IBD patients.
- Bifidobacterium infantis 35624 — the strain in Align. Originally developed for IBS, with subsequent observation of improvements in IBS-associated depression and anxiety.
The mechanisms by which psychobiotic strains produce mood effects involve all four gut-brain axis pathways: vagal afferent signaling (the Bravo mechanism), production of neurotransmitter precursors and SCFAs, modulation of HPA axis reactivity, and reduction of systemic inflammation through gut barrier reinforcement.
A frustrating limitation of the field is that "probiotic" effects are highly strain-specific. Lactobacillus rhamnosus JB-1 produces anxiolytic effects; other L. rhamnosus strains, including the closely related GG strain widely sold commercially, do not. Consumers buying "Lactobacillus" capsules generally cannot tell whether they are getting the strain that was actually studied for the indication they want to address. The Clinical Guide to Probiotic Products (clinicalguide.usprobioticguide.com) is a regularly-updated resource mapping documented indications to specific commercial products and strains.
GABA and Dopamine-Precursor Producing Bacteria
Beyond influencing host serotonin synthesis, certain gut bacteria produce neuroactive molecules directly. The most studied:
- GABA production — several Lactobacillus species (L. brevis, L. plantarum, L. paracasei) and Bifidobacterium dentium express glutamate decarboxylase (GAD) and convert glutamate to GABA. The bacterial GABA does not cross the blood-brain barrier directly, but it can activate enteric GABA-A receptors and produce vagal afferent signaling.
- Dopamine precursors — certain gut bacteria produce tyrosine and L-DOPA. Enterococcus faecalis tyrosine decarboxylase activity has clinical relevance: in Parkinson's patients taking levodopa, gut bacterial decarboxylation of levodopa to dopamine in the small intestine reduces the fraction reaching the brain, reducing drug efficacy. This is one reason carbidopa is co-administered.
- Norepinephrine — Escherichia coli and certain Saccharomyces yeast strains produce norepinephrine.
- Acetylcholine — Lactobacillus plantarum and certain other strains produce small amounts of acetylcholine.
- Histamine — multiple bacterial species produce histamine (relevant to histamine intolerance and SIBO presentations).
The clinical implication is that the microbiome contributes to a low-level "chemical noise floor" of neurotransmitter and neurotransmitter precursor signaling in the gut lumen and lamina propria. The signals are largely sensed via vagal afferents and enteric neurons rather than producing systemic neurotransmitter elevations. For more on the GABA mechanism specifically, see the GABA page.
Why SSRIs Cause Gut Side Effects
Selective serotonin reuptake inhibitors (SSRIs) are designed to block serotonin reuptake at brain synapses by inhibiting the serotonin transporter (SERT). The problem is that SERT is also abundantly expressed in the gut — it normally takes up serotonin released by enterochromaffin cells, terminating the local signal. SSRI inhibition of gut SERT therefore prolongs and amplifies enterochromaffin serotonin signaling in the gut, which produces:
- Nausea and vomiting — especially in the first weeks of SSRI treatment, via 5-HT3 receptor activation on vagal afferents (the same receptor blocked by ondansetron for chemotherapy-induced nausea)
- Diarrhea — via 5-HT4 receptor activation on enteric neurons and accelerated colonic transit
- Less commonly, constipation — particularly with paroxetine, which has stronger anticholinergic activity
- Loss of appetite — via central and peripheral 5-HT2C receptor activation
- GI bleeding risk — platelet serotonin is required for normal platelet aggregation, and SSRIs deplete platelet serotonin over weeks, producing a modest antiplatelet effect (clinically meaningful when combined with NSAIDs or anticoagulants)
The reason GI side effects typically improve over 2-4 weeks of SSRI therapy is partial downregulation of gut 5-HT3 receptors with sustained exposure. The reason SSRIs occasionally cause persistent GI dysmotility (chronic IBS-like symptoms in some patients) is incomplete adaptation.
The clinical implication: in patients with comorbid functional GI disorders and depression/anxiety, the choice of antidepressant matters. Tricyclic antidepressants (especially amitriptyline at low doses) are often preferred for IBS-D because anticholinergic effects slow gut transit. SSRIs are sometimes preferred for IBS-C for the opposite reason. Mirtazapine has the most favorable GI side-effect profile of the major classes and is sometimes chosen for patients particularly sensitive to GI effects. See our IBS page for more.
Fecal Microbiota Transplantation for Depression
Fecal microbiota transplantation (FMT) is the transfer of stool from a healthy donor into a recipient's gut, typically via capsule, colonoscopic infusion, or nasoenteric tube. FMT is FDA-cleared only for recurrent Clostridium difficile infection, but it has been studied in dozens of other indications, including depression.
The proof-of-concept evidence for FMT in depression comes from animal studies. Transferring stool from depressed human patients into germ-free mice produces depressive-like behaviors in the recipient mice (reduced sucrose preference, prolonged immobility in forced swim test); transferring stool from healthy human controls does not. This is one of the strongest causal demonstrations that the gut microbiome can produce mood effects, not merely correlate with them.
Human trials of FMT for depression are early-stage. The Kelly 2017 pilot study (4 patients with treatment-resistant depression who received open-label FMT) reported clinical improvement in 4 of 4 patients at 8 weeks. Subsequent small randomized trials show variable effects, generally favoring FMT over sham but with effect sizes that are modest and confidence intervals that are wide. The field is still working out donor selection, FMT preparation, dosing, and the role of patient-specific factors.
FMT for depression is not currently a recommended clinical intervention outside research settings. The far more practical and evidence-supported alternatives are dietary modification, prebiotic fiber, and specific psychobiotic strains, all discussed in detail on the Restoration Protocols page.
Clinical Implications for Mood Disorders
The gut-brain serotonin axis is real and mechanistically important, but the therapeutic translation is more nuanced than popular accounts suggest. Practical implications for patients with mood disorders:
- Diet is the largest-effect intervention — the SMILES trial (Jacka 2017, BMC Medicine) randomized adults with moderate-to-severe major depression to dietary counseling toward a Mediterranean pattern or social support. The dietary arm achieved a 32% remission rate vs. 8% in the control arm at 12 weeks. Effect size larger than most pharmaceutical trials.
- Psychobiotic supplementation produces modest effects — meta-analyses show small but consistent reductions in depression and anxiety scores with probiotic supplementation, with effect sizes around 0.2-0.4. Most effective: combination formulas (especially L. helveticus R0052 + B. longum R0175) and B. longum 1714. Duration: at least 4-8 weeks of daily use.
- Fermented foods may outperform probiotic capsules — the Sonnenburg 2021 Cell study found that a high-fermented-foods diet (yogurt, kefir, kimchi, sauerkraut, kombucha, fermented vegetables, fermented teas, 6 servings/day) increased microbiome diversity and reduced 19 inflammatory markers over 10 weeks. A parallel high-fiber arm did not produce the same effects in this short timeframe.
- Address gut barrier integrity — if leaky gut is contributing, anti-inflammatory dietary patterns plus targeted nutrients (L-glutamine, zinc carnosine, deglycyrrhizinated licorice) can reduce systemic inflammation and IDO induction.
- Psychobiotics do not replace standard treatment for severe depression — SSRIs, SNRIs, ECT, and ketamine remain primary interventions for moderate-to-severe major depressive disorder. Gut-brain interventions are best positioned as adjuncts and as standalone interventions for mild-to-moderate symptoms.
- Tryptophan supplementation is a separate question — oral tryptophan and 5-HTP can raise brain serotonin synthesis but carry serotonin syndrome risk if combined with SSRIs or MAO inhibitors. See L-Tryptophan page.
Key Research Papers
- Yano JM, Yu K, Donaldson GP, Shastri GG, Ann P, Ma L, Nagler CR, Ismagilov RF, Mazmanian SK, Hsiao EY (2015). Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 161(2):264-276. — Search PubMed
- Bravo JA, Forsythe P, Chew MV, Escaravage E, Savignac HM, Dinan TG, Bienenstock J, Cryan JF (2011). Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. PNAS 108(38):16050-16055. — PubMed: Bravo 2011
- Allen AP, Hutch W, Borre YE, Kennedy PJ, Temko A, Boylan G, Murphy E, Cryan JF, Dinan TG, Clarke G (2016). Bifidobacterium longum 1714 as a translational psychobiotic. Translational Psychiatry 6(11):e939. — Search PubMed
- Messaoudi M, Lalonde R, Violle N, et al. (2011). Assessment of psychotropic-like properties of a probiotic formulation (L. helveticus R0052 and B. longum R0175) in rats and human subjects. British Journal of Nutrition 105(5):755-764. — PubMed: Messaoudi 2011
- Jacka FN, O'Neil A, Opie R, et al. (2017). A randomised controlled trial of dietary improvement for adults with major depression (the SMILES trial). BMC Medicine 15(1):23. — PubMed: SMILES trial
- O'Mahony SM, Clarke G, Borre YE, Dinan TG, Cryan JF (2015). Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behavioural Brain Research 277:32-48. — PubMed: O'Mahony tryptophan
- Strandwitz P (2018). Neurotransmitter modulation by the gut microbiota. Brain Research 1693(Pt B):128-133. — PubMed: Strandwitz
- Kelly JR, Allen AP, Temko A, et al. (2017). Lost in translation? The potential psychobiotic Lactobacillus rhamnosus (JB-1) fails to modulate stress or cognitive performance in healthy male subjects. Brain, Behavior, and Immunity 61:50-59. — PubMed: Kelly translation
- Wastyk HC, Fragiadakis GK, Perelman D, et al. (2021). Gut-microbiota-targeted diets modulate human immune status. Cell 184(16):4137-4153. — PubMed: Sonnenburg fermented foods
- Valles-Colomer M, Falony G, Darzi Y, et al. (2019). The neuroactive potential of the human gut microbiota in quality of life and depression. Nature Microbiology 4(4):623-632. — PubMed: Valles-Colomer
- Maes M, Leonard BE, Myint AM, Kubera M, Verkerk R (2011). The new "5-HT" hypothesis of depression: cell-mediated immune activation induces indoleamine 2,3-dioxygenase. Progress in Neuro-Psychopharmacology & Biological Psychiatry 35(3):702-721. — Search PubMed
- Reigstad CS, Salmonson CE, Rainey JF 3rd, et al. (2015). Gut microbes promote colonic serotonin production through an effect of short-chain fatty acids on enterochromaffin cells. FASEB Journal 29(4):1395-1403. — PubMed: Reigstad SCFA
PubMed Topic Searches
- PubMed: Gut microbiota serotonin enterochromaffin
- PubMed: Psychobiotic depression anxiety
- PubMed: Kynurenine depression
- PubMed: Tryptophan gut-brain
- PubMed: SSRI GI side effects
Connections
- Gut-Brain Axis Hub
- Gut-Brain Benefits Deep-Dive
- Vagal Nerve Signaling
- Stress and Permeability
- Restoration Protocols
- L-Tryptophan
- GABA
- Depression
- Anxiety
- Irritable Bowel Syndrome
- Natural Anxiety Relief
- Stress Management
- Fermented Foods
- Probiotics
- All Remedies