Extended Fasts 24 to 72 Hours — Deep Autophagy & Stem-Cell Renewal
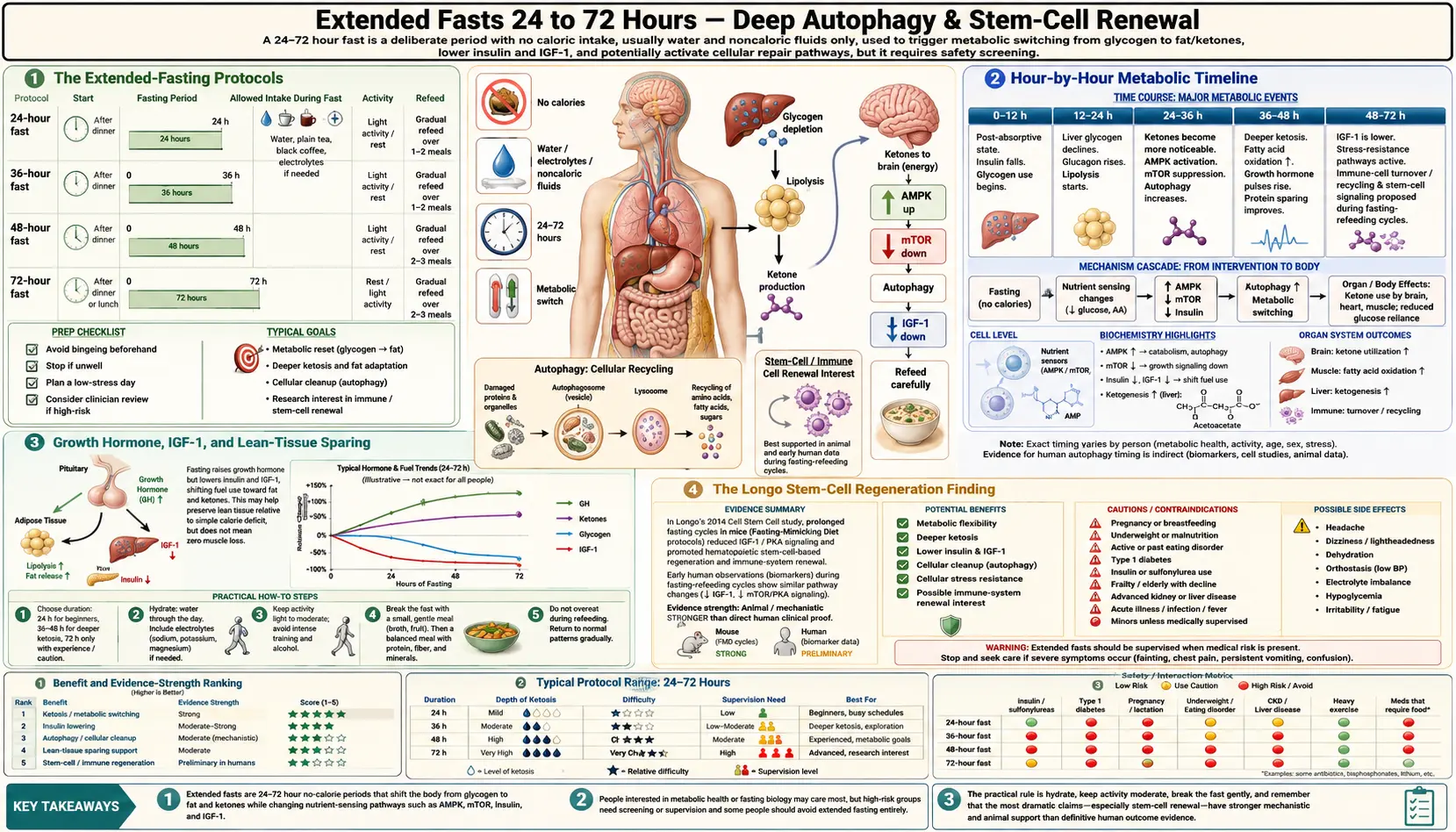
When the daily 16:8 protocol is the entry-level dose of fasting, extended fasts of 24, 36, 48, or 72 hours are where the dramatic metabolic, immune, and regenerative effects become measurable. Hepatic glycogen is exhausted by hour 18-24 in most adults, after which beta-hydroxybutyrate climbs steadily — reaching 1-2 mmol/L by 24 hours, 2-4 mmol/L by 48 hours, and 4-6 mmol/L by 72 hours. Growth hormone secretion increases 5-fold by 24 hours of fasting (Hartman et al.), an evolved mechanism to spare lean tissue while mobilizing fat. Valter Longo's 2014 Cell Stem Cell paper documented that 72-hour fasting cycles in mice and humans triggered apparent regeneration of the hematopoietic stem cell compartment — the bone-marrow precursors that produce all blood and immune cells. This page maps the dose-response curve from a single 24-hour skip to the 72-hour fast that defines the upper limit of self-directed at-home practice.
Interactive Visualization Fat Burning & Ketosis — what actually happens Follow a fatty acid out of a fat cell to the mitochondrial gate and find it padlocked by insulin. Drop the carbs, open CPT1, and watch the liver start making ketones. Launch →
Table of Contents
- The Extended-Fasting Protocols
- Hour-by-Hour Metabolic Timeline
- Growth Hormone, IGF-1, and Lean-Tissue Sparing
- The Longo Stem-Cell Regeneration Finding
- The Fasting-Mimicking Diet (FMD)
- Deep Ketosis and the BHB Signaling Cascade
- Cancer-Adjuvant Applications (Longo Differential Stress Resistance)
- Electrolyte and Hydration Protocol
- Cautions Specific to Extended Fasts
- Key Research Papers
- Connections
- Featured Videos
The Extended-Fasting Protocols
- 24-hour fast (Eat-Stop-Eat) — popularized by Brad Pilon. Eat normally on day 1, fast from dinner-to-dinner (e.g. 7 PM Monday to 7 PM Tuesday), then resume eating. Practiced 1-2 times per week. The lowest-friction entry point to extended fasting.
- 36-hour fast (Krista Varady ADF) — alternate-day fasting in its strict form: eat normally on day 1, complete fast on day 2, eat normally on day 3, fast on day 4. The modified version permits 500 calories on fasting days (the 5:2 diet popularized by Michael Mosley).
- 48-hour fast — the practical sweet spot for many practitioners: long enough to reach 2-3 mmol/L BHB, drive significant autophagy, and produce the 5-fold growth hormone surge; short enough to remain safely self-directed without medical supervision in healthy adults.
- 72-hour fast — the upper bound of unsupervised at-home extended fasting in most clinical recommendations. This is the duration used in Longo's stem-cell regeneration work. Beyond 72 hours, the risk-benefit balance typically requires supervision (a clinic like Buchinger Wilhelmi, or at minimum a physician familiar with prolonged fasting).
- 5-day prolonged fast or fasting-mimicking diet (FMD) — the standard duration used in Longo's clinical trials. The full water-only 5-day fast is generally inferior to the calorie-restricted FMD (described below) because the FMD captures most of the metabolic benefit with much better tolerability and safety.
Hour-by-Hour Metabolic Timeline
- Hours 0-4 (postprandial) — circulating glucose and amino acids from the last meal are the dominant fuels. Insulin is elevated. mTOR is active. Anabolic state.
- Hours 4-8 (early fast) — postprandial insulin declines. Hepatic glycogenolysis begins to supply blood glucose. mTOR begins to descend. AMPK begins to rise.
- Hours 8-12 — hepatic glycogen substantially depleted. Gluconeogenesis from lactate, glycerol, and glucogenic amino acids becomes the primary glucose source. Fat oxidation accelerates.
- Hours 12-18 (the metabolic switch) — ketone bodies begin to appear in measurable concentration (0.2-0.5 mmol/L BHB). Daily 16:8 practitioners typically reside in this zone.
- Hours 18-24 (entering moderate ketosis) — BHB climbs to 1-2 mmol/L in a metabolically flexible person, slower in someone insulin-resistant. Hunger peaks then plateaus as ghrelin oscillates and BHB suppresses appetite. Many practitioners report the hunger of hour 22-24 is the worst of any extended fast.
- Hours 24-36 (deepening ketosis, GH surge) — BHB approaches 2-3 mmol/L. Growth hormone pulses amplify dramatically (Hartman finding: ~5-fold increase by 24 hours). Lipolysis is maximal. Autophagy markers (LC3-II, autophagosome formation) increase substantially in animal models.
- Hours 36-48 — BHB 3-4 mmol/L. Mental clarity and energy often improve as the brain adapts to ketone fuel. Ghrelin hunger has typically peaked and declined. The fasting practitioner often feels paradoxically better at hour 40 than at hour 24.
- Hours 48-72 — BHB 4-6 mmol/L. Maximal autophagy in tissue studies. PKA pathway downregulation, IGF-1 decline. The Longo 72-hour stem-cell regeneration signal emerges in this window.
- Beyond 72 hours — diminishing returns on most metabolic markers, increasing risk of electrolyte derangement and refeeding-syndrome potential. Medical supervision warranted.
Growth Hormone, IGF-1, and Lean-Tissue Sparing
The intuitive concern about extended fasting is muscle loss. The empirical reality is more interesting: fasting produces a coordinated hormonal response that preserves lean tissue while preferentially mobilizing fat.
Hartman et al. demonstrated in healthy young men that growth hormone (GH) secretion increases approximately 5-fold during a 24-hour fast and remains elevated through 48-72 hours. GH stimulates lipolysis (release of free fatty acids from adipose tissue) and antagonizes insulin's effects on glucose disposal. The metabolic logic is straightforward: fasting elevates GH to mobilize fat for fuel and to suppress protein breakdown that would otherwise be needed to maintain blood glucose via gluconeogenesis.
Simultaneously, insulin-like growth factor 1 (IGF-1) declines during prolonged fasting — the opposite direction from GH. This apparent paradox reflects that fasted-state GH is acting primarily through its lipolytic and counter-regulatory effects, not its growth-promoting IGF-1-mediated effects. The decline in IGF-1 is one of the proposed mechanisms behind fasting's longevity signal — lower lifetime IGF-1 exposure is associated with reduced cancer incidence and extended lifespan in multiple animal models and human cohort observations.
The combination — high GH, low IGF-1, high lipolysis, suppressed insulin — produces a fuel-partition strategy that preserves muscle protein during reasonable-duration fasts. Cahill's classic 1970 NEJM work on prolonged starvation demonstrated that nitrogen excretion (a marker of protein breakdown) actually declines after the first 1-3 days of fasting as the body shifts increasingly to ketone fuel and reduces its glucose demand. The naive expectation that "you'll lose muscle if you don't eat protein every 4 hours" is not supported by the metabolic data for fasts up to 7 days in well-nourished adults.
The Longo Stem-Cell Regeneration Finding
Valter Longo and colleagues at USC Davis published a remarkable 2014 Cell Stem Cell paper (Cheng CW et al.) showing that prolonged fasting cycles — specifically 48-120 hours of water-only or FMD — produced apparent regeneration of the hematopoietic stem cell (HSC) compartment in the bone marrow of both mice and humans.
The mechanism: fasting reduces circulating IGF-1 and PKA (protein kinase A) signaling. Lower PKA in hematopoietic stem cells de-represses the transcription factor FOXO and reduces the activity of mTORC1. This combination appears to enable HSC self-renewal and produces a wave of new immune cell production during the refeeding phase after a prolonged fast.
Clinically the team demonstrated that cancer patients on chemotherapy who fasted 48-72 hours before treatment had improved immune-cell recovery profiles — suggesting that the stem-cell regeneration finding may have practical implications for protecting hematopoietic function during cytotoxic therapy. The work has not yet translated into routine clinical recommendations, but it is the basis for the active clinical trials of fasting-mimicking diet as a chemotherapy adjunct.
The implication for healthy practitioners: a single annual or quarterly 72-hour fast may produce a similar wave of immune-cell turnover — conceptually analogous to factory-rebooting the immune compartment. Mechanistic claims of "stem cell rejuvenation" require caution because the human data are limited and the long-term clinical outcomes are not yet established. But the mouse data are reproducible across multiple labs, and the biological plausibility is strong.
The Fasting-Mimicking Diet (FMD)
Recognizing that 5-day water-only fasts are difficult, often unsafe outside supervision, and require lengthy adaptation, the Longo lab developed the fasting-mimicking diet (FMD) — a 5-day low-calorie, low-protein, plant-based food regimen designed to produce the metabolic and hormonal signatures of fasting (low IGF-1, low glucose, elevated ketones, suppressed mTOR) while still providing some food intake to improve safety and tolerability.
The standard FMD protocol used in Longo's clinical trials:
- Day 1 — ~1100 calories: 11% protein, 46% fat, 43% carbohydrate. Plant-based.
- Days 2-5 — ~725 calories: 9% protein, 44% fat, 47% carbohydrate. Plant-based.
The commercial product (ProLon) packages this as soups, bars, and drinks. The DIY version is similar: vegetable broth, a small portion of nuts and olive oil, leafy greens, herbal teas, no animal protein, very limited carbohydrate.
Wei, Longo et al. (2017) randomized 100 generally healthy adults to three monthly cycles of the 5-day FMD versus control. The FMD group showed reductions in body weight, body fat, blood pressure, fasting glucose, IGF-1, and CRP — the entire metabolic-syndrome cluster. Critically, the benefits persisted at follow-up, suggesting the periodic FMD reset can produce durable improvement in cardiometabolic risk markers in people who otherwise eat normally between cycles.
The FMD is the practical translation of extended fasting for people who cannot tolerate or safely do a 5-day water-only fast — which is most people.
Deep Ketosis and the BHB Signaling Cascade
The 16:8 page introduced beta-hydroxybutyrate as a signaling molecule, not just a fuel. In the deeper ketosis of extended fasting (BHB 3-6 mmol/L), the signaling effects become substantially more pronounced:
- HDAC class I inhibition — at higher concentrations, BHB more effectively inhibits histone deacetylases, allowing histone acetylation and de-repressed expression of antioxidant genes (FOXO3, MT2, catalase, superoxide dismutase)
- NLRP3 inflammasome suppression — the Youm 2015 mechanism becomes more clinically relevant at the higher BHB levels of extended fasting
- GPR109A receptor activation — BHB activates the GPR109A G-protein-coupled receptor (the same receptor activated by niacin), which produces anti-lipolytic effects in adipose tissue and may contribute to the appetite suppression of deep ketosis
- BDNF and neuroprotection — the deeper ketosis of extended fasting more strongly upregulates BDNF in the hippocampus, the proposed mechanism for the cognitive clarity and mood elevation many extended-fast practitioners report by hour 36-48
- Mitochondrial biogenesis — ketosis upregulates PGC-1alpha, the master transcriptional coactivator for mitochondrial biogenesis, increasing mitochondrial density in tissues over repeated fasting cycles
The mitochondrial-biogenesis effect is one of the reasons practitioners often report sustained energy improvements over months of intermittent fasting practice — not just during the fast itself but in the fed state between fasts.
Cancer-Adjuvant Applications (Longo Differential Stress Resistance)
One of the more intriguing translational arms of Longo's work concerns fasting as an adjunct to cancer chemotherapy. The hypothesis — "differential stress resistance" (DSR) — is that normal cells, when starved of glucose and growth factors, downshift their metabolism, suppress mTOR, and become temporarily resistant to the toxic effects of chemotherapy. Cancer cells, with their characteristic constitutively-active growth signaling (oncogenic Ras, PI3K, mTOR), cannot downshift — they remain in growth mode regardless of the fasting signal, and are therefore differentially vulnerable to chemotherapy during the fasted state.
Raffaghello, Longo et al. demonstrated this differential vulnerability in cell-culture and mouse models. Multiple small clinical pilot trials in patients receiving platinum-based or anthracycline-based chemotherapy have shown that 48-72 hour pre-treatment fasting reduces patient-reported chemotherapy side effects (nausea, fatigue, weakness) and shows trends toward improved immune-cell recovery.
The clinical translation is still preliminary — this is not yet standard-of-care, and any patient considering fasting around chemotherapy must do so only in coordination with their oncologist and clinical nutritionist, with careful monitoring of nutritional status. But the mechanism is biologically sound and the early human data are encouraging enough that multiple Phase II and Phase III trials are now underway internationally.
Electrolyte and Hydration Protocol
The single most common reason extended fasts are abandoned prematurely is electrolyte deficiency — specifically sodium, magnesium, and potassium — manifesting as headache, fatigue, muscle cramps, palpitations, or orthostatic hypotension. These symptoms are often misattributed to "low blood sugar" but they are usually electrolyte-driven.
The reason: during the first 2-3 days of fasting, insulin falls substantially. Low insulin signals the kidneys to excrete sodium (the "natriuresis of fasting"). The lost sodium drags water and other electrolytes with it. Without active replacement, a fasting practitioner can lose 1-2 liters of fluid and several grams of sodium per day in urine alone.
The recommended daily electrolyte intake during an extended fast:
- Water: 2-3 liters, sipped throughout the day. Do not drink large boluses (causes dilutional symptoms).
- Sodium: 3-5 grams (1.5-2.5 tsp salt) — pinches of mineral salt dissolved in water across the day, or from broth.
- Potassium: 1-3 grams — "lite salt" (potassium chloride substitute) is the simplest source; vegetable broth provides smaller amounts.
- Magnesium: 300-500 mg — magnesium glycinate or citrate. Magnesium is the most commonly underdosed electrolyte in fasting practitioners and is the usual cause of leg cramps and poor sleep during extended fasts.
Commercial electrolyte powders designed for fasting (LMNT, Snake Juice, Dr. Berg's) package the right ratios. They contain zero calories and do not break the fast on any reasonable definition.
Cautions Specific to Extended Fasts
- All TRE cautions apply with greater force — pregnancy, lactation, children, T1 diabetes, eating disorder history, underweight, elderly with sarcopenia. Extended fasting is a more demanding intervention with smaller safety margins.
- Medication adjustment is essential — insulin, sulfonylureas, blood pressure medications (especially diuretics, ACE inhibitors, ARBs), and SGLT2 inhibitors all interact dangerously with extended fasting. Do not attempt without physician adjustment of doses.
- Gout — fasting can precipitate gout flares due to elevated uric acid (lactate and BHB share renal transporters with urate, blocking urate excretion). Patients with established gout should consult their physician and may need to continue allopurinol/febuxostat during the fast.
- Gallstones — rapid weight loss including from extended fasting can precipitate gallstone formation. Patients with known cholelithiasis should be cautious.
- Refeeding syndrome — the most serious complication of extended fasting, particularly after fasts > 5 days or in patients who began at low body weight. See the refeeding strategy page for the specific protocols to prevent it.
- Orthostatic hypotension — common in extended fasts, especially with inadequate sodium. Stand up slowly. Discontinue the fast if symptoms are severe.
- Beyond 72 hours — medical supervision is strongly recommended. Risk-benefit deteriorates without expert oversight.
Key Research Papers
- Cahill GF Jr. (1970). Starvation in man. NEJM. — PubMed
- Cheng CW et al. (Longo lab, 2014). Prolonged fasting reduces IGF-1/PKA to promote hematopoietic-stem-cell-based regeneration and reverse immunosuppression. Cell Stem Cell. — PubMed
- Wei M et al. (Longo lab, 2017). Fasting-mimicking diet and markers/risk factors for aging, diabetes, cancer, and cardiovascular disease. Science Translational Medicine. — PubMed
- Brandhorst S et al. (Longo lab, 2015). A periodic diet that mimics fasting promotes multi-system regeneration, enhanced cognitive performance, and healthspan. Cell Metabolism. — PubMed
- Hartman ML et al. Augmented growth hormone secretory burst frequency and amplitude during prolonged fasting in normal adults. JCEM. — PubMed
- Stekovic S et al. (2019). Alternate day fasting improves physiological and molecular markers of aging in healthy, non-obese humans. Cell Metabolism. — PubMed
- Trepanowski JF et al. (2017). Effect of alternate-day fasting on weight loss, weight maintenance, and cardioprotection. JAMA Internal Medicine. — PubMed
- Wilhelmi de Toledo F et al. (2019). Safety, health improvement, and well-being during a 4 to 21-day fasting period in an observational study including 1422 subjects. PLoS ONE. — PubMed
- Raffaghello L et al. (Longo lab, 2008). Starvation-dependent differential stress resistance protects normal but not cancer cells against high-dose chemotherapy. PNAS. — PubMed
- Bauersfeld SP et al. (2018). The effects of short-term fasting on quality of life and tolerance to chemotherapy in patients with breast and ovarian cancer. BMC Cancer. — PubMed
- Catenacci VA et al. (2016). A randomized pilot study comparing zero-calorie alternate-day fasting to daily caloric restriction in adults with obesity. Obesity. — PubMed
- Furmli S et al. (2018). Therapeutic use of intermittent fasting for people with type 2 diabetes as an alternative to insulin. BMJ Case Reports. — PubMed
PubMed Topic Searches
- PubMed: Prolonged fasting and autophagy
- PubMed: Fasting GH and IGF-1
- PubMed: Fasting-mimicking diet clinical
- PubMed: Alternate-day fasting
- PubMed: Fasting and chemotherapy
Connections
- Fat Burning & Ketosis — interactive animation
- Fasting Benefits Hub
- Fasting Overview
- Time-Restricted Eating 16:8
- Autophagy & mTOR
- Refeeding Strategy
- Type 2 Diabetes
- Obesity
- Metabolic Syndrome
- Fatty Liver Disease
- Oncology
- Magnesium (Fasting Electrolyte)
- Potassium (Fasting Electrolyte)
- Sodium (Fasting Electrolyte)
- Bone Broth
- All Remedies