Berberine for Gut Microbiome
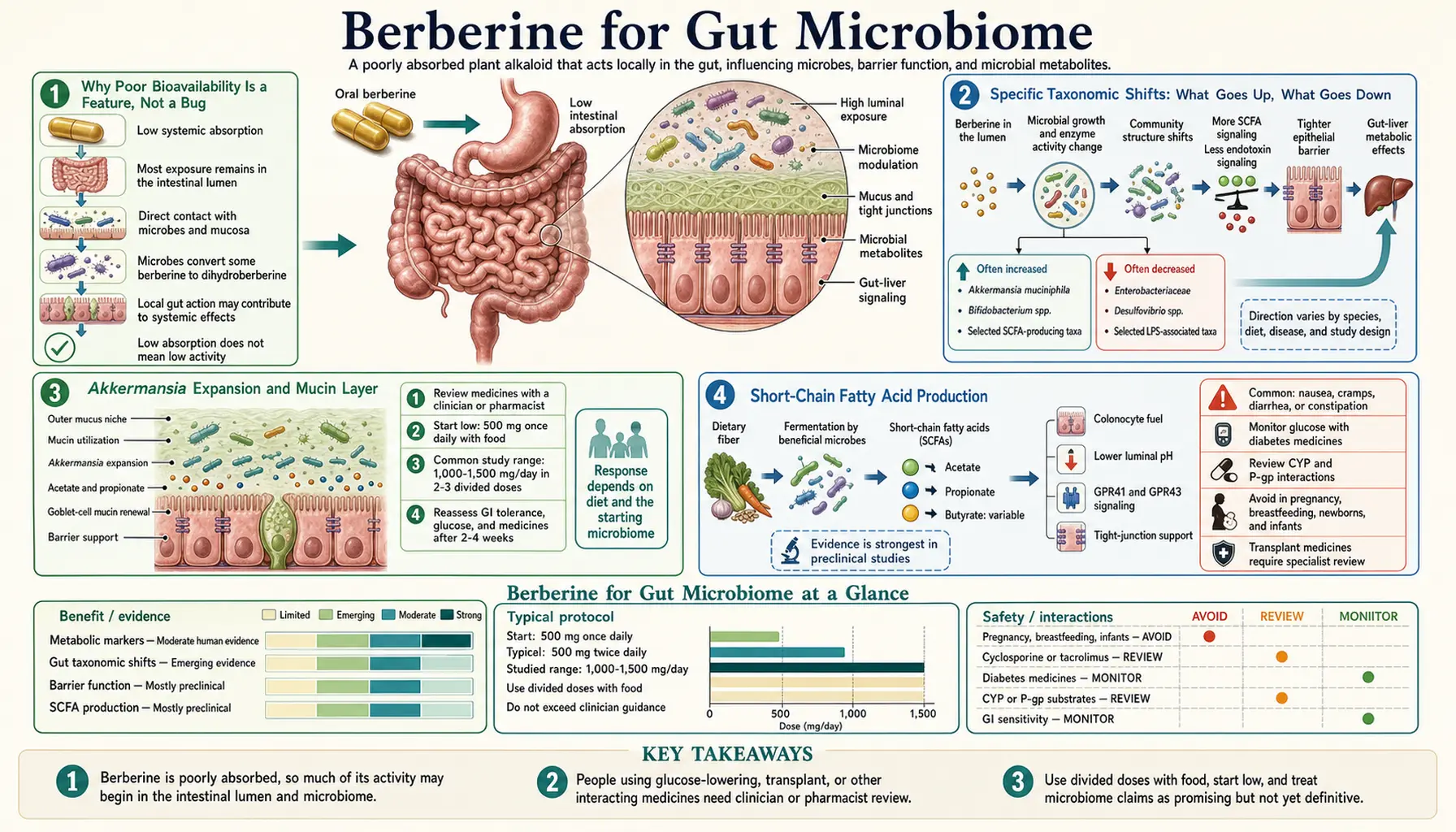
Berberine's 1-5% oral bioavailability used to be considered its main pharmacological problem — how could a molecule with such poor systemic absorption produce metformin-comparable effects on glucose and statin-comparable effects on lipids? The answer, increasingly clear over the past decade of microbiome research, is that the unabsorbed 95-99% of an oral berberine dose is not wasted. It is the active fraction for an entirely separate mechanism — gut microbiome remodeling. Berberine in the lower gastrointestinal lumen reaches concentrations far higher than anywhere else in the body, and at those concentrations it selectively suppresses lipopolysaccharide-producing Enterobacteriaceae and Gram-negative pathobionts while permitting expansion of short-chain-fatty-acid producers like Akkermansia muciniphila and Faecalibacterium prausnitzii. The downstream effects — reduced metabolic endotoxemia, improved gut barrier integrity, reduced systemic inflammation, secondary improvements in insulin sensitivity and lipid metabolism through the gut-liver-adipose axis — explain a meaningful fraction of berberine's clinical benefit. This page walks through the microbiome biology, the specific taxonomic shifts, the SIBO application, and the broader gut-liver-metabolic axis.
Table of Contents
- Why Poor Bioavailability Is a Feature, Not a Bug
- Specific Taxonomic Shifts: What Goes Up, What Goes Down
- Akkermansia Expansion and Mucin Layer
- Short-Chain Fatty Acid Production
- Lipopolysaccharide and Metabolic Endotoxemia
- Intestinal Barrier and Tight Junction Integrity
- Bile Acid Metabolism and FXR Signaling
- Small Intestinal Bacterial Overgrowth (SIBO)
- Historical Use as Antimicrobial and Antidiarrheal
- The Gut-Mediated Mechanism Behind Glucose and Lipid Effects
- Key Research Papers
- Connections
- Featured Videos
Why Poor Bioavailability Is a Feature, Not a Bug
The bioavailability story is worth telling carefully because it inverts the standard pharmacological intuition. For most drugs, low oral bioavailability is a problem to be solved — the desired effect is at a systemic target, and reaching it requires absorbed drug in circulation. Drug development typically focuses on improving absorption, formulating to bypass first-pass metabolism, increasing systemic exposure.
For berberine, the situation is fundamentally different. The systemic action (AMPK activation in liver, muscle, and adipose tissue) is real and clinically important — this is the basis for the metformin-comparable glucose effect documented in the trial data. But it is achieved with very low systemic concentrations because the molecular target (AMPK) is exquisitely sensitive, and even nanomolar tissue concentrations are sufficient.
Meanwhile, the unabsorbed 95-99% of the oral dose passes through the duodenum, jejunum, ileum, and colon at very high luminal concentrations — in the millimolar range, which is 1,000-fold higher than the systemic blood concentration. At these concentrations, berberine is a meaningfully potent antimicrobial with selective activity. It is not a broad-spectrum sterilizer like a fluoroquinolone — it has differential effects on different bacterial taxa, suppressing some while permitting others, in patterns that have turned out to be beneficial.
Studies tracking the fate of orally administered berberine have shown:
- Plasma berberine levels rarely exceed 5 ng/mL even with the 1,500 mg/day standard dose
- Luminal concentrations in the small intestine and colon reach 500-2000 micromolar (100,000-fold higher than plasma)
- A meaningful fraction is converted in the gut by bacterial nitroreductase to dihydroberberine, which has higher oral absorption and is then re-oxidized to berberine in the bloodstream — a "prodrug-like" relationship between the gut and the systemic compartment
- The remaining berberine is excreted in the feces along with conjugated metabolites
So berberine is genuinely two drugs in one: a systemic AMPK activator at low plasma concentrations, and a luminal gut microbiome modulator at high local concentrations. The clinical effect is the sum of both, and meta-analyses have not been able to fully partition which fraction of the glucose or lipid benefit comes from each route.
Specific Taxonomic Shifts: What Goes Up, What Goes Down
Multiple 16S rRNA sequencing studies in humans and animals have mapped the consistent microbiome shifts produced by oral berberine. The general pattern:
Taxa typically reduced:
- Enterobacteriaceae family (includes E. coli, Klebsiella, Enterobacter) — these Gram-negative pathobionts produce lipopolysaccharide and are increased in metabolic syndrome and type 2 diabetes
- Desulfovibrio — sulfate-reducing bacteria associated with inflammatory bowel disease and intestinal barrier disruption
- Proteobacteria phylum broadly — expansion of this phylum is a marker of dysbiosis
- Several Bacteroides species (the effect is species-specific, not phylum-wide)
- Ruminococcus gnavus — associated with inflammatory bowel disease and mucin degradation
Taxa typically expanded:
- Akkermansia muciniphila — a mucin-degrading bacterium strongly associated with metabolic health (covered separately below)
- Faecalibacterium prausnitzii — the principal butyrate-producing bacterium in the human colon, anti-inflammatory, depleted in IBD
- Lactobacillus species — the classic probiotic genus, beneficial for immune function and barrier integrity
- Bifidobacterium species — another classic probiotic genus, declines with age and is associated with multiple beneficial outcomes
- Roseburia — another butyrate-producer in the Lachnospiraceae family
The net shift — suppression of Gram-negative pathobionts and expansion of Gram-positive short-chain-fatty-acid producers — is approximately the opposite of the dysbiotic pattern seen in metabolic syndrome, type 2 diabetes, NAFLD, and inflammatory bowel disease. The conceptual framing is that berberine functions as a "microbiome corrector" — selectively suppressing the bacteria associated with metabolic dysfunction and expanding the bacteria associated with metabolic health.
This profile is broadly similar to the effects of metformin on the gut microbiome (which has its own well-documented effect on Akkermansia expansion), and the two are likely additive when combined.
Akkermansia Expansion and Mucin Layer
Akkermansia muciniphila deserves separate discussion because it has emerged in the past decade as perhaps the single most important "metabolically beneficial" bacterium in the human gut. Akkermansia is a Verrucomicrobia phylum organism that resides in the mucus layer overlying the colonic epithelium, where it specializes in degrading mucin (the glycoprotein component of intestinal mucus).
Counterintuitively, Akkermansia degradation of mucin is beneficial, not harmful. The mechanism is that Akkermansia's mucin degradation provides a signal to the colonic goblet cells to increase mucin production, leading to a thicker, more dynamic mucus layer over time. The mucus layer is the principal physical barrier separating the colonic epithelium from the bacterial mass in the lumen; thicker mucus = less bacterial translocation = less inflammatory signaling.
Akkermansia is depleted in obesity, type 2 diabetes, inflammatory bowel disease, and metabolic syndrome. Restoring Akkermansia has become a major research target, and several mechanisms accomplish it: metformin, dietary cranberry, dietary polyphenols, intermittent fasting, and oral berberine all expand Akkermansia in human trials.
The Zhang et al. 2012 paper in PLoS One was the first to systematically document berberine's Akkermansia-expanding effect. In high-fat-diet-fed rats, berberine 100 mg/kg/day produced a 90% reduction in body weight gain compared to high-fat controls, accompanied by ~10-fold expansion of Akkermansia abundance. Subsequent human studies have confirmed the directionality of the effect, though the magnitude in humans is more modest than in the rodent model.
The therapeutic significance is that berberine is an Akkermansia-expander that can be taken orally, at a known dose, with predictable pharmacology — in contrast to attempts to administer Akkermansia directly as a probiotic (which is technically difficult because Akkermansia is an anaerobe that does not survive standard probiotic processing well).
Short-Chain Fatty Acid Production
Short-chain fatty acids (SCFAs) — principally acetate, propionate, and butyrate — are produced by colonic bacterial fermentation of dietary fiber and resistant starch. They serve multiple roles:
- Butyrate is the principal energy substrate for colonocytes (colon epithelial cells get 60-70% of their energy from butyrate, not from circulation). Adequate butyrate maintains epithelial integrity and reduces inflammation.
- Propionate is absorbed and travels via portal circulation to the liver, where it suppresses hepatic gluconeogenesis and lipogenesis.
- Acetate circulates systemically and acts as a substrate for cholesterol synthesis (in some contexts) and as a signal through G-protein-coupled receptors (GPR41, GPR43) that modulate appetite, energy expenditure, and immune function.
SCFAs are decreased in metabolic syndrome and type 2 diabetes, both because the underlying microbiome has fewer SCFA-producing bacteria and because dietary fiber intake is often low. Berberine's expansion of Faecalibacterium, Roseburia, and related butyrate-producers translates into measurable increases in fecal and serum SCFA concentrations in supplementation trials.
The downstream metabolic benefits of restored SCFA production are substantial and partially explain berberine's glucose, lipid, and weight effects through pathways entirely independent of the systemic AMPK mechanism:
- Propionate-mediated suppression of hepatic gluconeogenesis (parallel to but mechanistically distinct from the direct AMPK effect on the liver)
- GLP-1 secretion from L-cells stimulated by SCFA signaling (parallel to the DPP-4 inhibition mechanism)
- PYY secretion contributing to satiety (the weight-loss effect of berberine in obese subjects)
- Anti-inflammatory effects mediated by butyrate's inhibition of histone deacetylase
Lipopolysaccharide and Metabolic Endotoxemia
Lipopolysaccharide (LPS), also called endotoxin, is a component of the outer membrane of Gram-negative bacteria. Low-level chronic translocation of LPS from the gut lumen into the portal circulation — "metabolic endotoxemia" — has emerged as a major driver of low-grade systemic inflammation in metabolic syndrome, type 2 diabetes, NAFLD, and obesity. LPS binds Toll-like receptor 4 (TLR4) on hepatocytes, Kupffer cells, adipocytes, and immune cells, triggering NF-kB activation and chronic inflammatory cytokine production (TNF-alpha, IL-6, IL-1-beta).
The mechanism of metabolic endotoxemia is twofold:
- Increased LPS production in the gut — from expansion of LPS-producing Gram-negative bacteria (Enterobacteriaceae, certain Bacteroides species), which is the dysbiotic pattern of metabolic syndrome
- Increased LPS translocation across the gut barrier — from impaired tight junction integrity (so-called "leaky gut")
Berberine addresses both. By suppressing Enterobacteriaceae, it reduces LPS production at the source. By improving tight junction integrity (the mechanism discussed below), it reduces LPS translocation. The net effect is a measurable reduction in serum LPS levels in supplementation trials — typically 20-30% reduction over 3 months — with corresponding reductions in serum CRP, TNF-alpha, and IL-6.
For patients with NAFLD, this gut-liver axis is particularly relevant. The "gut-liver-fat" model of NAFLD progression centers on LPS-driven hepatic inflammation as the trigger that converts simple hepatic steatosis (NAFL) to nonalcoholic steatohepatitis (NASH), which is the histologically inflamed precursor to cirrhosis. Berberine's gut-LPS-reducing effect adds to its direct hepatic AMPK effect to produce the substantial benefit documented in NAFLD trials. See our NAFLD page for the broader management discussion.
Intestinal Barrier and Tight Junction Integrity
The intestinal epithelial barrier is maintained by specialized intercellular junctions: tight junctions (most apically located, formed by claudins, occludin, ZO-1, and JAM-A), adherens junctions, desmosomes, and gap junctions. These junctions regulate paracellular permeability — the leakiness of the gut wall to molecules in the lumen.
In states of intestinal inflammation (IBD, celiac, SIBO) or metabolic stress, tight junction proteins are reduced or disorganized, leading to increased paracellular permeability and the translocation of bacterial products (LPS, peptidoglycan, flagellin) and food antigens into circulation. This is the molecular basis of "leaky gut."
Berberine produces measurable improvements in tight junction integrity:
- Increased expression of ZO-1 (zonula occludens-1), occludin, and claudin-1 in colonic epithelium
- Reduced expression of claudin-2 (a "leaky" claudin that creates paracellular pores)
- Reduced transepithelial flux of FITC-dextran (a standard permeability marker) in animal models
- Reduced serum zonulin (a marker of intestinal permeability) in human studies
The mechanism appears to be a combination of direct effects on epithelial cells (some AMPK-mediated, some independent) and indirect effects through the SCFA-producing bacteria whose products (especially butyrate) are themselves potent inducers of tight junction protein expression.
The therapeutic implication is that berberine is one of the more reliable agents for repairing impaired intestinal barrier function. Other agents with similar effects include L-glutamine, zinc carnosine, slippery elm, and certain probiotics; berberine has the advantage of additional metabolic benefits beyond the barrier effect alone. For more on barrier integrity, see our Leaky Gut page.
Bile Acid Metabolism and FXR Signaling
Bile acids are not just digestive surfactants — they are also signaling molecules that bind nuclear receptors (principally farnesoid X receptor, FXR) and membrane G-protein-coupled receptors (TGR5), regulating glucose, lipid, and energy metabolism. Bile acid signaling has emerged as one of the major mechanisms connecting the gut microbiome to systemic metabolic health, because gut bacteria perform critical transformations of primary bile acids (cholic acid, chenodeoxycholic acid) to secondary bile acids (deoxycholic acid, lithocholic acid) with different receptor affinities.
Berberine modulates bile acid pools in several ways:
- Direct binding to FXR (described in some studies, contested in others — the affinity is modest if real)
- Modulation of bile-acid-deconjugating bacteria (bile salt hydrolase activity), shifting the conjugated/unconjugated ratio
- Modulation of bile-acid-7-alpha-dehydroxylating bacteria, shifting the primary/secondary ratio
- Indirect effects through FXR signaling on hepatic cholesterol metabolism, gluconeogenesis, and triglyceride synthesis
The clinical relevance is that bile acid signaling is now a major drug target — obeticholic acid (an FXR agonist) is approved for primary biliary cholangitis, and is in trials for NASH. Berberine's modulation of the bile acid pool is one more pathway connecting it to NAFLD/NASH benefit, and there is ongoing research on whether berberine's FXR-related effects contribute meaningfully to its overall metabolic benefit.
Small Intestinal Bacterial Overgrowth (SIBO)
Small intestinal bacterial overgrowth (SIBO) is the inappropriate expansion of bacteria in the small intestine, where bacterial counts should be 1000-fold lower than in the colon. SIBO produces bloating, abdominal pain, diarrhea or constipation, malabsorption, and is increasingly recognized as a contributor to irritable bowel syndrome, fibromyalgia, and rosacea among other conditions.
Standard SIBO therapy is the antibiotic rifaximin (550 mg three times daily for 14 days), which is poorly absorbed and works locally in the small intestine. Berberine has emerged as a credible non-antibiotic alternative based on:
- The Chedid 2014 study in Global Advances in Health and Medicine compared a herbal protocol containing berberine to rifaximin in 104 patients with SIBO. Response rates were comparable: 46% for the herbal protocol versus 34% for rifaximin.
- Berberine's broad-spectrum antimicrobial activity at high luminal concentrations, with documented activity against the principal SIBO organisms (E. coli, Klebsiella, Streptococcus, Staphylococcus, Bacteroides)
- The motility-promoting effect of berberine on the migrating motor complex, which is the principal physiological defense against SIBO (impaired motility is the underlying driver of most SIBO cases)
- The bonus systemic benefits (glucose, lipids) that are absent with rifaximin
Typical SIBO protocols using berberine include berberine 500 mg three times daily for 4-6 weeks, often combined with another antimicrobial herb (oregano oil, neem, allicin) and a prokinetic agent (low-dose erythromycin, prucalopride, ginger) to prevent relapse. The relapse rate without ongoing prokinetic support is approximately 40% within 6 months, comparable to rifaximin's relapse rate.
See our SIBO page and SIBO prokinetics deep dive for the broader management discussion.
Historical Use as Antimicrobial and Antidiarrheal
Berberine's modern reputation as a metabolic agent is relatively recent (since approximately 2004). Its historical use, by contrast, was almost entirely as a gastrointestinal antimicrobial and antidiarrheal — in traditional Chinese medicine (Huang Lian, from Coptis chinensis), Ayurveda (from Berberis aristata), and Western herbalism (from Hydrastis canadensis, goldenseal).
The historical indications — infectious diarrhea, dysentery, cholera-like illness, parasitic infection — align with the modern understanding of berberine as a luminal antimicrobial. Controlled trials in the 1980s and 1990s established efficacy of oral berberine for:
- Cholera-associated diarrhea (mechanism: inhibition of intestinal secretion driven by cholera toxin's effect on adenylyl cyclase)
- Enterotoxigenic E. coli diarrhea (parallel mechanism)
- Giardiasis (modest efficacy; metronidazole more effective)
- Acute infectious gastroenteritis broadly
The mechanism of antimicrobial action at high luminal concentrations involves multiple targets: DNA intercalation, FtsZ inhibition (disrupting bacterial cell division), efflux pump inhibition (which can re-sensitize antibiotic-resistant bacteria), and inhibition of secretory toxins. The combination of mechanisms makes resistance development less likely than with single-target antibiotics — though for the modern uses of berberine (chronic metabolic indications), the question of antimicrobial resistance is less acute because the relevant target is the host metabolism, not pathogen eradication.
The Gut-Mediated Mechanism Behind Glucose and Lipid Effects
It is worth synthesizing the gut effects with the systemic effects covered on the other Benefits pages, because the integration explains how a molecule with 1-5% oral bioavailability produces metformin-comparable systemic effects.
The proposed integrated model:
- Oral berberine reaches the small intestine and colon at high luminal concentrations
- Berberine selectively remodels the microbiome — suppressing LPS-producing Gram-negative pathobionts, expanding SCFA-producing Gram-positives and Akkermansia
- The remodeled microbiome reduces metabolic endotoxemia (less LPS translocation) and increases SCFA production (more butyrate, propionate, acetate)
- Reduced LPS-driven inflammation in the liver and adipose tissue improves insulin sensitivity (TNF-alpha is a known driver of insulin resistance through serine phosphorylation of IRS-1)
- Propionate suppresses hepatic gluconeogenesis through portal circulation, adding to the direct AMPK effect on the liver
- SCFA-mediated GLP-1 release amplifies postprandial insulin secretion and reduces glucagon, lowering postprandial glucose
- Improved tight junction integrity reduces ongoing antigen translocation, breaking the feed-forward inflammatory cycle
- Meanwhile, the small absorbed fraction reaches the liver, muscle, and adipose tissue and produces the direct AMPK-mediated effects on glucose disposal, lipogenesis, and gluconeogenesis covered on the Blood Sugar deep-dive page
The sum of the systemic AMPK effect plus the gut-mediated effects is what produces the metformin-comparable clinical outcome. Either mechanism alone would be modest; the combination is substantial.
Key Research Papers
- Zhang X, Zhao Y, Zhang M et al. (2012). Structural changes of gut microbiota during berberine-mediated prevention of obesity and insulin resistance in high-fat diet-fed rats. PLoS One, 7(8):e42529. — PubMed 22880019
- Xie W, Gu D, Li J et al. (2011). Effects and action mechanisms of berberine and Rhizoma coptidis on gut microbes and obesity in high-fat diet-fed rats. PLoS One, 6(9):e24520. — PubMed 21915347
- Wang Y, Shou JW, Li XY et al. (2017). Berberine-induced bioactive metabolites of the gut microbiota improve energy metabolism. Metabolism, 70:72-84. — PubMed 28403947
- Cao Y, Pan Q, Cai W et al. (2016). Modulation of gut microbiota by berberine improves steatohepatitis in high-fat diet-fed BALB/C mice. Archives of Iranian Medicine, 19(3):197-203. — Search PubMed
- Habtemariam S (2020). Berberine pharmacology and the gut microbiota: A hidden therapeutic link. Pharmacological Research, 155:104722. — PubMed 32105754
- Chedid V, Dhalla S, Clarke JO et al. (2014). Herbal therapy is equivalent to rifaximin for the treatment of small intestinal bacterial overgrowth. Global Advances in Health and Medicine, 3(3):16-24. — PubMed 24891990
- Feng R, Shou JW, Zhao ZX et al. (2015). Transforming berberine into its intestine-absorbable form by the gut microbiota. Scientific Reports, 5:12155. — PubMed 26174047
- Sun H, Wang N, Cang Z et al. (2016). Modulation of microbiota-gut-brain axis by berberine resulting in improved metabolic status in high-fat diet-fed rats. Obesity Facts, 9(6):365-378. — Search PubMed
- Gu S, Cao B, Sun R et al. (2015). A metabolomic and pharmacokinetic study on the mechanism underlying the lipid-lowering effect of orally administered berberine. Molecular BioSystems, 11(2):463-474. — Search PubMed
- Tan XS, Ma JY, Feng R et al. (2013). Tissue distribution of berberine and its metabolites after oral administration in rats. PLoS One, 8(10):e77969. — Search PubMed
- Liu Y, Hao H, Xie H et al. (2010). Extensive intestinal first-pass elimination and predominant hepatic distribution of berberine explain its low plasma levels in rats. Drug Metabolism and Disposition, 38(10):1779-1784. — PubMed 20634337
- Cui HX, Hu YN, Li JW et al. (2018). A purified anthraquinone-glycoside preparation from Rheum officinale inhibits Helicobacter pylori in vitro and protects against ethanol-induced gastric ulcer in vivo — comparison with berberine. Frontiers in Pharmacology, 9:1067. — Search PubMed
PubMed Topic Searches
- PubMed: Berberine gut microbiome
- PubMed: Berberine Akkermansia
- PubMed: Berberine LPS endotoxemia
- PubMed: Berberine SIBO
- PubMed: Berberine tight junction barrier
Connections
- Berberine Overview
- Berberine Benefits Hub
- Berberine for Blood Sugar
- Berberine for Cholesterol
- Berberine Dosing & Cautions
- SIBO
- SIBO Prokinetics
- Leaky Gut
- Irritable Bowel Syndrome
- NAFLD
- Oregano Oil
- Allicin
- Resistant Starch
- Probiotics