Walnuts: ALA Omega-3 and Brain Health
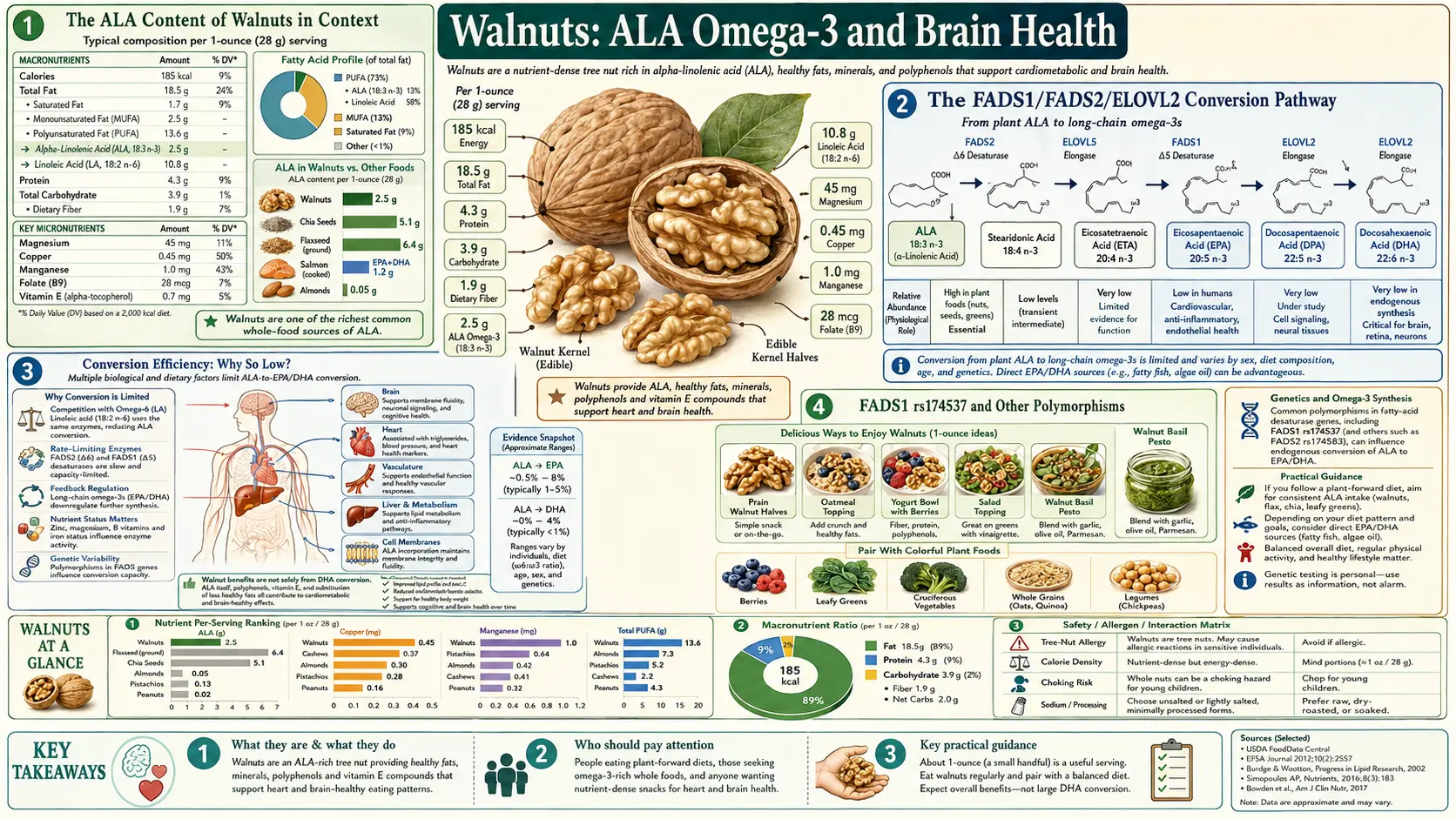
A single ounce of walnuts delivers 2.5 grams of alpha-linolenic acid (ALA) — more than the entire daily Adequate Intake of plant omega-3 for an adult, and more than any other commonly consumed nut. But ALA is not the same molecule as the EPA and DHA that the brain actually uses for membrane phospholipids and inflammatory signaling. The human body must elongate and desaturate ALA through the FADS1/FADS2/ELOVL2 enzymatic pathway to produce EPA and DHA, and that conversion is famously inefficient — typically 5-10% of ingested ALA reaches EPA and less than 1% reaches DHA in healthy adults. This deep-dive walks through the biochemistry, the genetic variants that determine whether someone is an efficient or inefficient converter, the direct brain effects of ALA that are independent of DHA conversion, and the practical question of whether walnuts can replace fatty fish or whether they complement it.
Table of Contents
- The ALA Content of Walnuts in Context
- The FADS1/FADS2/ELOVL2 Conversion Pathway
- Conversion Efficiency: Why So Low?
- FADS1 rs174537 and Other Polymorphisms
- Sex Differences: Women Convert Better Than Men
- Direct Brain Effects of ALA Independent of DHA
- Brain Phospholipid Incorporation and Hippocampal Volume
- Walnut Versus Fish: Replacement or Complement?
- Practical Dosing and Combining Strategies
- Cautions
- Key Research Papers
- Connections
- Featured Videos
The ALA Content of Walnuts in Context
Alpha-linolenic acid (ALA, 18:3 n-3) is the parent compound of the omega-3 family. It is a polyunsaturated fatty acid with eighteen carbons and three double bonds, the first of which sits at the third carbon from the methyl end — the structural feature that defines an omega-3. ALA is an essential fatty acid: the human body cannot synthesize it de novo and must obtain it from the diet.
Walnuts are exceptional among nuts in their ALA content. Per 100 grams (about three and a half ounces):
- Walnuts (English): 9.1 g ALA — the standout
- Flaxseed (ground): 22.8 g ALA — higher, but a seed not a nut
- Chia seed: 17.8 g ALA
- Hemp seed (hulled): 8.7 g ALA
- Pecans: 1.0 g ALA
- Pistachios: 0.3 g ALA
- Almonds, cashews, hazelnuts, macadamias, brazils, pine nuts: essentially 0 g ALA
Per one-ounce (28 g) serving — the typical "handful of nuts" portion — walnuts deliver about 2.5 g ALA. The Institute of Medicine Adequate Intake (AI) for ALA is 1.6 g/day for adult men and 1.1 g/day for adult women. A single one-ounce serving of walnuts exceeds the daily AI by 50-130%. No other widely consumed nut comes anywhere close.
This is why walnuts occupy a unique position in any conversation about plant omega-3 intake. Among foods that people actually eat in normal serving sizes, only fatty fish, flaxseed, chia, and walnuts deliver meaningful omega-3. Of those four, walnuts are the most convenient to eat as a snack or recipe ingredient.
The FADS1/FADS2/ELOVL2 Conversion Pathway
ALA is not the omega-3 molecule that brain tissue uses preferentially. The brain incorporates docosahexaenoic acid (DHA, 22:6 n-3) into its membrane phospholipids — DHA accounts for approximately 15% of total brain phospholipid fatty acids and is particularly concentrated in synaptic membranes and the photoreceptor outer segments of the retina. Eicosapentaenoic acid (EPA, 20:5 n-3) plays a separate role in resolving neuroinflammation through specialized pro-resolving mediators like resolvin E1.
To get from dietary ALA to brain DHA, the human body uses a multistep elongation and desaturation pathway:
- ALA (18:3 n-3) → SDA (18:4 n-3) — catalyzed by FADS2 (delta-6 desaturase), the rate-limiting enzyme of the pathway
- SDA (18:4 n-3) → ETA (20:4 n-3) — catalyzed by ELOVL5 (elongase)
- ETA (20:4 n-3) → EPA (20:5 n-3) — catalyzed by FADS1 (delta-5 desaturase)
- EPA (20:5 n-3) → DPA (22:5 n-3) — catalyzed by ELOVL2/ELOVL5
- DPA (22:5 n-3) → tetracosapentaenoic acid (24:5 n-3) — ELOVL2
- Tetracosapentaenoic (24:5 n-3) → tetracosahexaenoic (24:6 n-3) — FADS2 again
- Peroxisomal beta-oxidation shortens 24:6 to DHA (22:6 n-3) — "Sprecher pathway"
This is a long, multistep, energy-requiring pathway that consumes the same enzymes the body uses for converting omega-6 linoleic acid to arachidonic acid, creating direct substrate-level competition between the omega-6 and omega-3 series. In a high-omega-6 Western diet (typical omega-6 to omega-3 ratio 15:1 to 25:1), the omega-6 substrate flood largely outcompetes ALA for the available FADS2/FADS1 enzyme activity, further reducing the ALA-to-DHA conversion efficiency.
Conversion Efficiency: Why So Low?
The classic studies (Burdge and Calder, Brenna, Williams, and others using isotope-labeled ALA tracer studies) have established conversion efficiencies that are sobering for anyone hoping plant omega-3 alone will meet brain DHA needs:
- ALA to EPA: approximately 5-10% in healthy adult men, slightly higher in women
- ALA to DPA (the intermediate): approximately 2-5%
- ALA to DHA: approximately 0.5-9%, with most studies finding the figure in the 1-2% range in men and 4-9% in women of reproductive age
To put this in absolute terms: if you eat one ounce of walnuts and consume 2.5 g of ALA, you might get 125-250 mg of EPA and 12-50 mg of DHA delivered to your tissues. Compare to a 100-gram serving of farmed salmon, which delivers approximately 1.5-2.5 g of EPA+DHA directly with no conversion needed.
The reasons for low conversion efficiency are multiple and largely structural:
- FADS2 is rate-limiting and saturable. The enzyme has finite throughput and is also used by the omega-6 pathway for arachidonic acid synthesis.
- Substrate competition with omega-6. A typical Western diet provides 15-25 times more linoleic acid (omega-6) than ALA. The competition for FADS2 favors the more-abundant substrate.
- The Sprecher pathway is metabolically expensive. The final ALA-to-DHA step requires elongation past the desired chain length, retroconversion via peroxisomal beta-oxidation, and inter-organelle trafficking — an unusual and energy-costly arrangement.
- Beta-oxidation of ALA for energy is a major competing fate. A substantial fraction of dietary ALA is simply burned for ATP rather than elongated to longer-chain omega-3s.
- DHA itself feedback-inhibits the conversion enzymes. When DHA status is already adequate, conversion downregulates — which is appropriate physiology but means people with low baseline DHA see modest increases that plateau.
FADS1 rs174537 and Other Polymorphisms
Conversion efficiency is not uniform across the population — it depends substantially on common single nucleotide polymorphisms (SNPs) in the FADS gene cluster on chromosome 11. The most-studied variant is rs174537, located in the FADS1 promoter region. This SNP has two common alleles:
- G allele (major in European ancestry, ~65% frequency): associated with higher FADS1 expression and more efficient conversion of ALA to EPA and DHA
- T allele (minor in European ancestry, ~35% frequency, but the major allele in some African populations): associated with lower FADS1 expression and less efficient conversion
The functional consequence is real and measurable. Homozygous GG individuals can convert several-fold more ALA to long-chain omega-3 than TT individuals fed the same dietary ALA. This polymorphism (and the related rs174448, rs1535, rs174556 cluster) accounts for a substantial fraction of inter-individual variation in serum and tissue EPA/DHA concentrations independent of dietary intake.
There is also a fascinating population-genetics layer: the high-converting GG haplotype shows signs of strong positive selection in populations historically dependent on plant-source diets (parts of South Asia, the Inuit before contact, agricultural populations from the Neolithic transition onward), while populations historically dependent on marine omega-3 (some coastal hunter-gatherer groups) show retention of the ancestral TT-converting low-efficiency variants — presumably because conversion was unnecessary when DHA was directly available from seafood. This is an example of gene-diet coevolution.
For the individual contemplating walnut versus fish-oil supplementation, the practical implication is that genetic conversion ability matters. Direct-to-consumer genetic testing (23andMe, Ancestry data run through a tool like Promethease) can give you your rs174537 status. If you are GG (high converter), plant omega-3 sources may serve you reasonably well. If you are TT (low converter), supplementing with preformed EPA/DHA from algal or fish oil is more likely to be necessary.
Sex Differences: Women Convert Better Than Men
One of the most consistent findings in the ALA-conversion literature is a sex difference favoring women, particularly women of reproductive age. Burdge's 2002 studies in British men and women found ALA-to-DHA conversion rates of approximately 9% in women versus less than 1% in men.
The mechanism is thought to be estrogen-driven upregulation of FADS2 expression. Estradiol increases hepatic delta-6 desaturase activity, presumably as an evolutionary adaptation to support the high DHA demands of fetal brain development during pregnancy and lactation. The DHA content of breast milk is approximately 0.3% of total fat and depends substantially on maternal long-chain omega-3 status — a measurable burden on maternal stores during the lactation period.
The practical implications:
- Premenopausal women may receive proportionally more brain-relevant DHA from a given ALA intake than men of the same age
- Postmenopausal women lose the estrogen-driven conversion advantage and become more similar to men in conversion efficiency
- Pregnant and lactating women have substantially elevated DHA needs that ALA alone rarely meets; preformed DHA supplementation (algal or fish oil) is generally recommended
- Hormonal contraceptives that contain estrogen may slightly increase conversion efficiency, though the clinical relevance is modest
For more on omega-3 needs across the life cycle, see our Omega-3 Fatty Acids page.
Direct Brain Effects of ALA Independent of DHA
The focus on ALA-to-DHA conversion can obscure a separate question: does ALA itself have brain-relevant effects independent of conversion? The answer is increasingly clear that it does, through at least three mechanisms.
- PPAR-alpha activation. ALA is a direct ligand for peroxisome proliferator-activated receptor alpha (PPAR-alpha), a nuclear transcription factor expressed in brain microglia and astrocytes. PPAR-alpha activation reduces production of pro-inflammatory cytokines (TNF-alpha, IL-6, IL-1-beta) and shifts microglial polarization toward an M2 anti-inflammatory phenotype. This is a non-conversion mechanism for ALA-driven neuroinflammation reduction.
- Direct incorporation into brain phospholipids. ALA itself is incorporated into brain phosphatidylcholine and phosphatidylethanolamine, though at much lower abundance than DHA. The local conversion of ALA to DHA within the brain (the brain has limited but real FADS2/FADS1 expression in astrocytes) provides a slow but steady supply of DHA independent of peripheral conversion.
- Substrate for brain-derived oxylipins. ALA is enzymatically oxidized to a family of bioactive lipid mediators including 9- and 13-HODE analogs of the omega-3 series, some of which have signaling roles in neuronal survival and synaptic plasticity.
The walnut clinical trials — PREDIMED-NAVARRA cognitive substudy, WAHA, Pribis 2012 inferential reasoning, observational cohorts on cognitive decline — consistently find some benefit to walnut consumption even though the ALA-to-DHA conversion is inefficient. This suggests that the direct-brain-effect mechanisms above contribute meaningfully alongside whatever DHA elevation is achieved by conversion. See the Cognitive Aging deep-dive for the clinical evidence detail.
Brain Phospholipid Incorporation and Hippocampal Volume
A subset of human imaging studies has examined whether walnut or ALA intake correlates with measurable brain structural changes. The findings are suggestive though not yet definitive.
Animal model work (Chauhan et al., Muthaiyah et al.) using walnut-extract supplementation in mouse models of Alzheimer's disease has demonstrated reductions in hippocampal amyloid-beta plaque burden, increased BDNF (brain-derived neurotrophic factor) expression in hippocampal tissue, and preserved memory performance on Morris water maze and Y-maze testing. The mechanism is proposed to involve both the ALA-omega-3 effect on membrane fluidity and the walnut polyphenol effect on amyloid aggregation kinetics.
In humans, the WAHA trial included an MRI substudy in a subset of participants that examined hippocampal and cortical volume changes over the two-year intervention period. The walnut arm showed numerically less hippocampal volume loss than the control arm, though the effect size was small and the comparison did not reach statistical significance in the full cohort — results were stronger in the Barcelona site than the Loma Linda site, raising questions about background diet effects.
The Rotterdam Study and the Three-City Study (both large European observational cohorts) have reported associations between higher dietary ALA intake and lower risk of cognitive decline and dementia at 5-10 year follow-up. These are not randomized trials, but they provide population-level signal that is consistent with the trial-level data on walnuts.
Walnut Versus Fish: Replacement or Complement?
The question that matters most for individual dietary planning is whether walnuts can replace fatty fish as an omega-3 source, or whether they should be regarded as a complement to fish.
The honest answer for most adults is "complement, not replacement." Here is the reasoning:
- For people with high FADS1 conversion (rs174537 GG) and adequate baseline omega-6:omega-3 ratio (achieved by avoiding seed oils and emphasizing whole foods), walnuts can deliver enough effective long-chain omega-3 to meet typical adult needs. This is probably about a third of the population.
- For low converters (rs174537 TT), people with elevated omega-6 intake, or people with elevated DHA demands (pregnancy, lactation, elderly with cognitive concerns, athletes with high inflammatory load), walnuts alone are unlikely to meet long-chain omega-3 needs. Adding fatty fish 2-3 times per week or supplementing with algal/fish oil is recommended.
- Walnuts and fatty fish are not redundant — they deliver overlapping but distinct nutrient packages. Walnuts add polyphenols (ellagitannins, urolithin precursors), fiber, magnesium, and copper. Fatty fish adds preformed EPA/DHA, complete protein, vitamin D, selenium, and iodine. Both belong in a varied diet.
For vegetarians and vegans who do not eat fish, the practical recommendation is daily walnuts (or flaxseed/chia) plus an algal DHA supplement (250-500 mg/day). Algal oil is the original source of marine DHA — fish are concentrators of algae-produced DHA via the food chain, and direct algal-oil supplementation bypasses the need for fish.
For omnivores: 2-3 fatty fish servings per week (salmon, sardines, mackerel, herring — small fatty fish low on the food chain to minimize mercury) plus daily walnuts gives a robust combination.
Practical Dosing and Combining Strategies
Practical recommendations for using walnuts as part of a brain-targeted nutrition strategy:
- Serving size: 1 ounce (28 g, about 14 walnut halves) per day. This delivers 2.5 g ALA, 185 calories, 18.5 g fat, 4 g protein, 2 g fiber, and the polyphenol package. Two ounces is reasonable for adults with elevated needs.
- Whole walnut vs walnut oil: Whole walnut is preferred because it preserves the polyphenol-rich pellicle (the brown papery skin contains most of the ellagitannins). Walnut oil delivers ALA but loses the polyphenols and most of the fiber.
- Form: Halves, pieces, or chopped — equivalent. Lightly toasted walnuts retain most of the ALA but lose some of the heat-sensitive polyphenols; raw or refrigerated is optimal.
- Storage: Walnuts are oxidation-prone because of the high ALA content. Refrigerate or freeze opened packages; do not store at room temperature for more than a month. Rancid walnuts taste bitter and have lost most of their omega-3 value.
- Timing: No clear time-of-day effect on conversion or brain incorporation. The convenient practice is to incorporate them into breakfast oatmeal, mid-morning snack, or evening salad.
- Pairing: The complete brain-omega-3 strategy: daily walnuts (ALA + polyphenols), 2-3x weekly fatty fish (preformed EPA/DHA), reduced seed-oil intake (lower omega-6 substrate competition), and adequate dietary magnesium (cofactor for FADS2 activity).
For people with documented low DHA status (Omega-3 Index test < 4%, available through OmegaQuant), walnuts alone will not correct the deficit in a reasonable time frame. Algal oil or fish oil supplementation at 1-2 g EPA+DHA per day for 3-6 months is typically required, followed by maintenance through dietary fatty fish.
Cautions
- Tree-nut allergy. Walnut is among the most common tree-nut allergens. Jug r 1 (2S albumin) and Jug r 3 (non-specific lipid transfer protein) are the major IgE-binding proteins. Walnut allergy can be severe and is sometimes the first tree-nut allergy to emerge in childhood. People with established tree-nut allergy should avoid walnuts entirely.
- Oxidative rancidity. The high ALA content makes walnuts unusually prone to oxidation. Rancid walnuts are bitter and have lost the omega-3 value. Refrigerate after opening; do not eat walnuts that smell or taste off.
- Aflatoxin contamination. Like other tree nuts, walnuts can be contaminated with aflatoxin from Aspergillus flavus mold if improperly dried or stored. Buy from reputable suppliers, store dry and refrigerated, and discard any walnut that shows mold or off-odor.
- Anticoagulant interaction. Walnut omega-3s have modest antiplatelet effects. Patients on warfarin or other anticoagulants should not dramatically increase walnut intake without informing their prescriber, though normal dietary intake (1-2 ounces daily) is generally not clinically significant.
- Calorie density. One ounce of walnuts is 185 calories. People monitoring weight should account for the calorie contribution; substituting walnuts for other snack foods is usually weight-neutral but adding them on top of an unchanged diet adds calories.
- Phytic acid and mineral absorption. See the Soaking and Phytates deep-dive. The phytate content is unlikely to cause mineral deficiency in adults with otherwise mixed diets, but can be a consideration in mineral-deficient populations.
Key Research Papers
- Burdge GC, Calder PC (2005). Conversion of alpha-linolenic acid to longer-chain polyunsaturated fatty acids in human adults. Reproduction, Nutrition, Development. — PubMed
- Burdge GC, Jones AE, Wootton SA (2002). Eicosapentaenoic and docosapentaenoic acids are the principal products of alpha-linolenic acid metabolism in young men. British Journal of Nutrition. — PubMed
- Burdge GC, Wootton SA (2002). Conversion of alpha-linolenic acid to eicosapentaenoic, docosapentaenoic and docosahexaenoic acids in young women. British Journal of Nutrition. — PubMed
- Brenna JT (2002). Efficiency of conversion of alpha-linolenic acid to long chain n-3 fatty acids in man. Current Opinion in Clinical Nutrition and Metabolic Care. — PubMed
- Schaeffer L et al. (2006). Common genetic variants of the FADS1 FADS2 gene cluster and their reconstructed haplotypes are associated with the fatty acid composition in phospholipids. Human Molecular Genetics. — PubMed
- Chilton FH et al. (2014). Diet-gene interactions and PUFA metabolism: a potential contributor to health disparities and human diseases. Nutrients. — PubMed
- Mathias RA et al. (2014). The impact of FADS genetic variants on omega-6 polyunsaturated fatty acid metabolism in African Americans. BMC Genetics. — PubMed
- Sprecher H (2000). Metabolism of highly unsaturated n-3 and n-6 fatty acids. Biochimica et Biophysica Acta. — PubMed
- Muthaiyah B et al. (2014). Dietary supplementation of walnuts improves memory and learning skills through mitigation of oxidative stress in Alzheimer's disease model mice. Journal of Alzheimer's Disease. — PubMed
- Chauhan A et al. (2014). Beneficial effects of walnuts on cognition and brain health. Nutrients. — PubMed
- Domenichiello AF, Kitson AP, Bazinet RP (2015). Is docosahexaenoic acid synthesis from alpha-linolenic acid sufficient to supply the adult brain? Progress in Lipid Research. — PubMed
- Bowen KJ et al. (2019). Omega-3 fatty acids and cardiovascular disease: are there benefits? Current Treatment Options in Cardiovascular Medicine. — PubMed
PubMed Topic Searches
- PubMed: Walnut ALA and brain
- PubMed: FADS1 rs174537 conversion
- PubMed: ALA PPAR-alpha neuroinflammation
- PubMed: Walnut and BDNF
- PubMed: Algal DHA supplementation
Connections
- Walnuts Overview
- Walnut Benefits Hub
- Walnuts and Cognitive Aging
- Walnuts and Heart Health (PREDIMED)
- Walnut Soaking and Phytates
- Omega-3 Fatty Acids
- Salmon
- Sardines
- Herring
- Neurology
- Vitamin E (Antioxidant Pair)
- Magnesium (FADS2 Cofactor)
- Mediterranean Diet
- Lab Tests (Omega-3 Index)
- All Food