PQQ for Mitochondrial Biogenesis
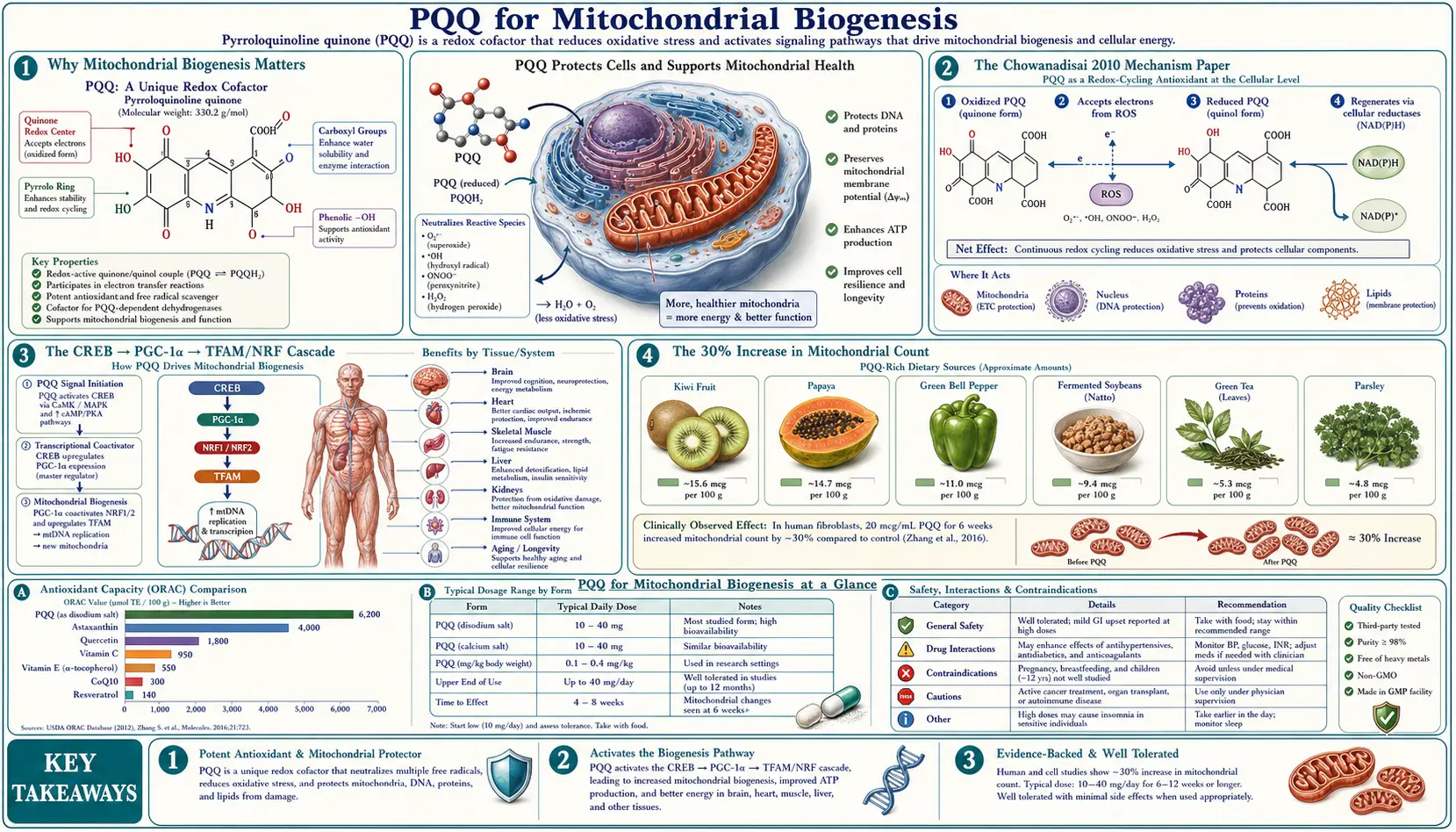
Mitochondrial biogenesis — the creation of new mitochondria within existing cells — is the single biochemical mechanism that separates PQQ from every other nutritional antioxidant. The Chowanadisai 2010 Journal of Biological Chemistry paper established that PQQ activates CREB phosphorylation, which upregulates PGC-1α expression, which in turn drives TFAM, NRF-1, and NRF-2 to assemble new mitochondria. Mouse liver shows a 30% increase in mitochondrial number per cell after 8 weeks of supplementation. This deep-dive walks through the mechanism step by step, contrasts PQQ with CoQ10 and ALA (number vs. function), explains why this is the "exercise-mimic without exercise" of the antioxidant world, and unpacks the implications for mitochondrial aging.
Table of Contents
- Why Mitochondrial Biogenesis Matters
- The Chowanadisai 2010 Mechanism Paper
- The CREB → PGC-1α → TFAM/NRF Cascade
- The 30% Increase in Mitochondrial Count
- Number vs. Function: PQQ vs. CoQ10 vs. ALA
- PQQ as an Exercise-Mimic Signal
- Contrast With Caloric Restriction
- Why This Matters for Mitochondrial Aging
- Practical Protocol & Timeline
- Cautions
- Key Research Papers
- Connections
- Featured Videos
Why Mitochondrial Biogenesis Matters
Mitochondrial density — the number of mitochondria per cell — declines progressively with age across most tissues. Skeletal muscle loses an estimated 5-10% of mitochondrial volume per decade after age 30. Brain tissue, particularly cortical neurons, shows similar decline. The heart, liver, and oocytes follow the same pattern. This loss is a major contributor to the "energy slowdown" of aging — reduced ATP synthesis capacity, reduced fat oxidation, reduced thermogenesis, and increased reactive oxygen species (ROS) leak from the smaller, more damaged remaining mitochondria.
Most mitochondrial supplements address this problem on the "function" side — they help existing mitochondria work better. CoQ10 feeds the electron transport chain. Alpha lipoic acid acts as cofactor for TCA-cycle enzymes. Methylene blue provides an alternative electron acceptor when complex damage is severe. Riboflavin rebuilds FAD/FMN cofactors for Complex I and II.
None of these grow new mitochondria. They all work within the existing complement of mitochondria the cell already has. PQQ is fundamentally different: it pharmacologically engages the master genetic switch that builds new mitochondria. This is the same switch flipped by aerobic exercise, caloric restriction, and cold exposure — all known biogenesis triggers. PQQ flips it through a nutritional pathway that doesn't require those physical stressors.
For most other supplements, you are upgrading a small factory. For PQQ, you are building a bigger factory. The practical implication is that PQQ pairs naturally with the function-enhancers rather than competing with them — the canonical PQQ + CoQ10 combination addresses both the number and the function of mitochondria simultaneously.
The Chowanadisai 2010 Mechanism Paper
The pivotal mechanistic paper is Chowanadisai W, Bauerly KA, Tchaparian E, Wong A, Cortopassi GA, Rucker RB. "Pyrroloquinoline quinone stimulates mitochondrial biogenesis through cAMP response element-binding protein phosphorylation and increased PGC-1α expression." Journal of Biological Chemistry 285(1):142-152, January 2010.
The UC Davis group started with a simple observation that had been floating around in PQQ literature for years: PQQ-deficient mice had fewer mitochondria per cell than PQQ-replete mice. The question was: how does a small organic cofactor regulate the assembly of an entire organelle?
Their experimental approach worked at three levels:
- In vivo (whole mouse): mice fed standard chow vs. PQQ-deficient chow vs. PQQ-supplemented chow for 8 weeks, then tissue analysis
- Ex vivo (isolated tissues): liver, heart, and skeletal muscle were assayed for mitochondrial DNA content, mitochondrial respiratory chain protein abundance, and gene expression of biogenesis regulators
- In vitro (cell culture): hepatocytes treated with physiologically relevant PQQ concentrations were analyzed for phosphorylation of upstream signaling molecules (CREB, AMPK) and induction of biogenesis transcription factors
The findings were unambiguous:
- Tissue mitochondrial DNA copy number was significantly higher in PQQ-supplemented vs. control mice
- Respiratory chain protein abundance (Complexes I, III, IV) increased in parallel
- Gene expression of PGC-1α, TFAM, NRF-1, and NRF-2 was upregulated 2-3-fold
- CREB phosphorylation in hepatocytes increased within hours of PQQ exposure
- Blocking CREB activation (with dominant-negative CREB constructs) abolished the downstream PGC-1α response
The paper established the full upstream-to-downstream signaling axis and has been cited several thousand times. Subsequent groups (Bauerly 2011, others) have replicated the core finding in multiple cell types and tissues.
The CREB → PGC-1α → TFAM/NRF Cascade
The signaling cascade that PQQ engages is the canonical mammalian mitochondrial-biogenesis pathway. The same cascade is engaged by exercise (via AMPK and calcineurin), by caloric restriction (via SIRT1), and by cold exposure (via β3-adrenergic signaling). PQQ converges on the pathway at the CREB step.
- PQQ → CREB phosphorylation. CREB (cAMP response element-binding protein) is a transcription factor activated by phosphorylation at serine 133. The Chowanadisai work showed PQQ rapidly increases CREB-Ser133 phosphorylation in hepatocytes. The proximal kinase responsible is most consistent with PKA activation downstream of an unidentified PQQ-engaged receptor, though the exact membrane target remains under investigation.
- Phospho-CREB → PGC-1α gene transcription. The PGC-1α promoter contains multiple CRE (cAMP response element) sites. Phospho-CREB binding to these elements drives transcription of the PGC-1α gene. Within 6-24 hours of PQQ exposure, PGC-1α mRNA increases 2-3-fold.
- PGC-1α → coactivation of nuclear respiratory factors. PGC-1α (peroxisome proliferator-activated receptor gamma coactivator 1-alpha) is the master transcriptional coactivator of mitochondrial biogenesis. It physically interacts with and coactivates NRF-1 and NRF-2 (nuclear respiratory factors 1 and 2), which bind to and activate the promoters of nuclear-encoded mitochondrial genes.
- NRF-1/NRF-2 → TFAM expression. TFAM (mitochondrial transcription factor A) is encoded in the nucleus but functions inside the mitochondrion, where it binds and packages mitochondrial DNA and drives transcription of the 13 mitochondrially encoded electron transport chain proteins. NRF-1 binding to the TFAM promoter drives the increase in TFAM that ultimately enables new mitochondrial DNA replication and respiratory chain assembly.
- TFAM → mitochondrial DNA replication. With increased TFAM available inside mitochondria, mitochondrial DNA is replicated, packaged into nucleoids, and used to template new respiratory chain assemblies. Combined with the increased supply of nuclear-encoded subunits from the NRF-1/NRF-2 program, the cell assembles physical new mitochondria over the following days.
The full cycle — from PQQ exposure to new mitochondrial assembly — takes roughly 4-7 days in cell culture and 2-8 weeks in tissues studied in vivo. This timeline matches the clinical-trial observation that PQQ effects on cognition, sleep, and fatigue typically take 8-12 weeks to become measurable.
The 30% Increase in Mitochondrial Count
The headline number from the Chowanadisai work is a ~30% increase in mitochondrial DNA copy number per cell in mouse liver after 8 weeks of PQQ supplementation at doses scaled to roughly correspond to 10-30 mg/day in humans. Heart and skeletal muscle showed smaller but similar-direction increases.
To put this in context:
- A typical hepatocyte has 1,000-2,000 mitochondria. A 30% increase means 300-600 additional mitochondria per cell.
- This is roughly equivalent to the mitochondrial biogenesis response seen after 6-12 weeks of moderate endurance training in skeletal muscle, though in a different tissue (liver).
- The increase reverses gradually after PQQ withdrawal — mitochondria turn over with a half-life of approximately 2-4 weeks, so the supplemental effect requires ongoing dosing to maintain.
Crucially, the new mitochondria are functional — respiratory chain protein abundance increased in proportion, suggesting the new mitochondria contribute to total ATP synthesis capacity rather than being structurally present but inert. This is meaningful because not all biogenesis stimuli produce equally functional mitochondria. Some pharmacological biogenesis activators (e.g., certain β3-agonists) produce mitochondria with incomplete respiratory chains; PQQ does not appear to have that defect.
Human studies have not directly measured mitochondrial copy number in solid tissue (this requires invasive biopsy) but have shown:
- Increased PGC-1α mRNA in peripheral blood mononuclear cells after PQQ supplementation
- Reduced markers of oxidative stress and inflammation (Harris 2013) consistent with improved mitochondrial function
- Improvements in cognitive and fatigue endpoints (Itoh, Nakano, Hwang) on a timeline consistent with the cell-culture and animal mechanism data
Number vs. Function: PQQ vs. CoQ10 vs. ALA
The single most useful conceptual frame for understanding the mitochondrial-nutrient space is the number vs. function distinction:
| Compound | Primary Mechanism | Number or Function? |
|---|---|---|
| PQQ | CREB → PGC-1α → new mitochondria | Number (biogenesis) |
| CoQ10 (ubiquinol) | Electron carrier between Complexes I/II and III | Function |
| Alpha lipoic acid | Cofactor for PDH, α-KGDH (TCA-cycle enzymes) | Function |
| Methylene blue | Alternative electron acceptor / Complex bypass | Function (rescue) |
| Riboflavin (B2) | FAD/FMN precursor for Complex I and II | Function (cofactor) |
| NAD+ / NMN / NR | NAD+ precursor, sirtuin activation, indirect PGC-1α effects | Mixed (some biogenesis via SIRT1) |
| Acetyl-L-carnitine | Fatty acid transport into mitochondria | Function |
| Creatine | ATP buffering via phosphocreatine | Function (storage) |
The asymmetry is striking. PQQ is essentially the only readily available oral compound that lives clearly on the "number" side. NAD+ precursors (NMN, NR) have some biogenesis effect through sirtuin activation but it is indirect and less robust than PQQ's direct CREB activation. Everything else is function.
This explains why the PQQ + CoQ10 combination has become so commercially successful and so clinically rational. The two compounds are doing genuinely complementary things: PQQ adds mitochondria; CoQ10 makes the new (and existing) mitochondria run more efficiently. Combined effects on biomarkers and clinical endpoints are additive.
PQQ as an Exercise-Mimic Signal
The PGC-1α cascade that PQQ engages is the same one engaged by aerobic exercise. In skeletal muscle, contractile activity activates AMPK (low-energy signaling) and calcineurin (calcium signaling), both of which converge on PGC-1α activation. The result is the well-documented mitochondrial biogenesis response to endurance training: ~30-50% increase in skeletal muscle mitochondrial volume after 8-12 weeks of moderate aerobic training.
This makes PQQ part of a small family of "exercise-mimic" compounds that pharmacologically activate biogenesis pathways. Other members of this category include:
- AICAR (5-aminoimidazole-4-carboxamide ribonucleoside) — direct AMPK activator, research compound only
- Metformin — AMPK activator, FDA-approved drug for diabetes, has shown longevity-extending effects in some animal models
- Resveratrol — SIRT1 activator (controversial mechanism), indirect biogenesis effects
- GW501516 (a synthetic PPAR-δ agonist) — works through a different transcription factor; banned in athletic competition due to cancer signals in animal studies
PQQ is essentially the only safe, well-tolerated, orally bioavailable, naturally occurring exercise-mimic compound. The clinical-trial evidence is more modest than for actual exercise — biogenesis from training in skeletal muscle is larger than biogenesis from oral PQQ — but the two are not mutually exclusive. PQQ + exercise produces additive biogenesis responses in studies that have measured both.
The practical clinical interpretation: PQQ is not a substitute for exercise. Aerobic activity remains the single most powerful biogenesis stimulus available, and the cardiovascular, metabolic, and psychiatric benefits of exercise extend far beyond mitochondrial number. But for populations where exercise is impossible or severely limited — the bedbound elderly, patients with severe heart failure or COPD, post-stroke patients with motor disability, ME/CFS patients with post-exertional malaise — pharmacological biogenesis activation via PQQ provides a partial substitute that exercise-based recommendations cannot.
Contrast With Caloric Restriction
Caloric restriction (CR) — 20-30% reduction in caloric intake while maintaining nutritional adequacy — is the most robust longevity intervention demonstrated across multiple species (yeast, worms, flies, mice, rats, rhesus monkeys, and observationally in humans). One of its core mechanisms is mitochondrial biogenesis via SIRT1 deacetylation of PGC-1α, which renders PGC-1α more active as a coactivator.
PQQ engages essentially the same downstream effector (PGC-1α) but via a different upstream signal (CREB phosphorylation rather than SIRT1-mediated PGC-1α deacetylation). The two mechanisms are mechanistically parallel and converge on the same biogenesis output.
Differences:
- CR has many additional benefits beyond biogenesis — reduced IGF-1 signaling, increased autophagy, improved insulin sensitivity, reduced inflammation, lower body composition. PQQ provides only the biogenesis arm of this multi-output response.
- CR is difficult to sustain. Adherence is the primary failure mode of CR as a clinical intervention. PQQ requires only daily pill-taking.
- CR-mimic compounds (rapamycin, metformin, NAD+ precursors) attempt to engage other CR mechanisms pharmacologically. PQQ engages the biogenesis arm. Combinations are biologically rational.
- CR + PQQ is not additive on biogenesis — both work through PGC-1α and converge on the same effector. The biogenesis response saturates around the maximum CR or PQQ effect alone.
For users interested in longevity-focused supplementation, the most defensible combination is PQQ (biogenesis) + a CR-mimic for the other arms (autophagy via spermidine or fasting-mimetic diets; sirtuin support via NAD+ precursors; mTOR modulation via intermittent low-protein periods).
Why This Matters for Mitochondrial Aging
The "mitochondrial theory of aging," articulated most influentially by Denham Harman in the 1970s and refined by many groups since, holds that the accumulation of mitochondrial damage — mtDNA mutations, respiratory chain inefficiency, increased ROS production, declining biogenesis — is a primary driver of organismal aging. Tissues most dependent on mitochondrial ATP (brain, heart, skeletal muscle, oocytes) are the tissues most affected by aging.
Within this framework, PQQ addresses one of the central mechanisms: the age-related decline in mitochondrial biogenesis. Tissues from old animals show measurably reduced PGC-1α activation in response to standard biogenesis stimuli (exercise, cold, fasting). Whether this is cause or consequence is debated, but the practical implication is the same: pharmacologically restoring biogenesis capacity should partially compensate.
The clinical conditions where this matters most include:
- Sarcopenia — age-related muscle loss is partly driven by mitochondrial biogenesis decline in skeletal muscle. PQQ + resistance training is mechanistically rational, though direct sarcopenia trials are limited.
- Cardiac aging and heart failure with preserved ejection fraction (HFpEF) — cardiomyocyte mitochondrial dysfunction is increasingly recognized in HFpEF. PQQ + CoQ10 is a reasonable adjunct to standard heart failure care.
- Cognitive aging — cortical neuron mitochondrial density falls with age. This is the mechanism most directly addressed by the cognitive trials. See the Cognition deep-dive.
- Oocyte aging and reduced IVF success — oocyte mitochondrial dysfunction is a major contributor to declining fertility with age. PQQ + CoQ10 is increasingly used in pre-IVF preparation, though robust trial data are limited.
- Acquired mitochondrial dysfunction syndromes — long-COVID, post-EBV reactivation, chronic Lyme, fibromyalgia, ME/CFS. The mitochondrial dysfunction in these conditions is heterogeneous, but biogenesis support is reasonable in conjunction with other mitochondrial nutrients.
Practical Protocol & Timeline
Standard biogenesis-focused protocol
- Dose: 20 mg/day BioPQQ or equivalent disodium salt
- Timing: Morning with breakfast (mild stimulant effect; evening dosing can disturb sleep)
- Paired with: Ubiquinol 100-200 mg/day (the function-side complement)
- Duration: 8-12 weeks minimum before assessing response; indefinite for longevity protocols
Week-by-week timeline
- Week 1-2: No perceptible effect; mild "more alert in the morning" sensation in some users (CREB activation is acute, but new mitochondrial assembly takes weeks)
- Week 3-4: Subjective energy and mental clarity often begin to improve. Sleep quality may improve subtly.
- Week 5-8: Steady accumulation of biogenesis effect. Cognitive measures (if testing) begin to show improvement.
- Week 8-12: Trial-endpoint window. This is when the published cognition trials measured their primary outcomes.
- Month 3-6: Plateau of biogenesis effect. Further benefit becomes incremental rather than dramatic. Maintenance phase begins.
- Year 1+: Stable mitochondrial biogenesis support as part of longevity stack.
Combinations
- PQQ + ubiquinol — the canonical pairing. 20 mg PQQ + 100-200 mg ubiquinol daily. Addresses both number and function.
- PQQ + alpha lipoic acid + acetyl-L-carnitine — the "Bruce Ames mitochondrial stack" with biogenesis layered on top. 20 mg PQQ + 600 mg ALA + 1000-2000 mg acetyl-L-carnitine.
- PQQ + NMN / NR — biogenesis + sirtuin activation. 20 mg PQQ + 250-500 mg NMN. Rational for longevity-focused users.
- PQQ + creatine — biogenesis + ATP buffering. Popular for cognitive performance and aging-related muscle loss.
Cautions
- Morning dosing for sleep protection — PQQ has a mild stimulant effect (consistent with improved mitochondrial energy output). Evening dosing can interfere with sleep onset in sensitive users, even though long-term PQQ improves sleep quality. Always take in morning or early afternoon.
- Headache (rare) — ~5-10% of users report mild headache during the first 1-2 weeks at 20-40 mg/day. Usually resolves with continued use or temporary dose reduction.
- Pregnancy — PQQ is naturally present in breast milk and dietary sources, but supplemental dosing during pregnancy lacks formal safety trials. Avoid in the absence of clinical guidance.
- Bleeding disorders — no documented anticoagulant effect; theoretical concern based on quinone chemistry only. Mention to physicians if you take warfarin or DOACs.
- Quality concerns — PQQ raw material is expensive; under-dosed and adulterated products are common. Prefer BioPQQ-labeled products or established brands with third-party testing.
- Realistic expectations — biogenesis-driven effects develop slowly over 8-12 weeks. Users expecting an acute energy boost (the kind delivered by caffeine or B-vitamins) will be disappointed. PQQ is a long-term mitochondrial intervention, not a short-term stimulant.
Key Research Papers
- Chowanadisai W et al. (2010). Pyrroloquinoline quinone stimulates mitochondrial biogenesis through cAMP response element-binding protein phosphorylation and increased PGC-1α expression. J Biol Chem 285(1):142-152. — PubMed: Chowanadisai 2010
- Bauerly K et al. (2011). Altered pyrroloquinoline quinone status alters mitochondrial respiratory chain function. PLoS One. — PubMed: Bauerly 2011
- Harris CB et al. (2013). Dietary pyrroloquinoline quinone (PQQ) alters indicators of inflammation and mitochondrial-related metabolism in human subjects. J Nutr Biochem. — PubMed: Harris 2013
- Stites T et al. (2006). Pyrroloquinoline quinone modulates mitochondrial quantity and function in mice. J Nutr. — PubMed: Stites 2006
- PGC-1α as master regulator of mitochondrial biogenesis (Wu et al.) — PubMed: Wu PGC-1α master regulator
- TFAM in mitochondrial DNA replication and transcription — PubMed: TFAM mtDNA replication
- NRF-1 and NRF-2 nuclear respiratory factors — PubMed: NRF-1 / NRF-2
- Exercise-induced PGC-1α expression in skeletal muscle — PubMed: exercise PGC-1α skeletal muscle
- Caloric restriction, SIRT1, and PGC-1α deacetylation — PubMed: CR / SIRT1 / PGC-1α
- Mitochondrial density decline in aging skeletal muscle — PubMed: mitochondrial density aging muscle
- PQQ in cardiac ischemia-reperfusion (cardiomyocyte biogenesis) — PubMed: PQQ cardiac ischemia
- PQQ in oocyte mitochondrial quality and IVF preparation — PubMed: PQQ oocyte / IVF
PubMed Topic Searches
- PubMed: PQQ mitochondrial biogenesis
- PubMed: PQQ PGC-1α
- PubMed: PQQ CREB phosphorylation
- PubMed: PQQ TFAM / NRF
- PubMed: PGC-1α aging decline
Connections
- PQQ Overview
- PQQ Benefits Hub
- PQQ for Cognition
- PQQ for Sleep & Mood
- PQQ for NGF Synthesis
- CoQ10 (function-side pairing)
- ALA for Mitochondria
- Methylene Blue
- NAD+ & NMN
- All Antioxidants
- Oxidative Stress
- Longevity Protocols
- Creatine
- Vitamin B2 (Riboflavin)
- Vitamin B3 (Niacin / NAD+ precursor)
- Fatigue
- Brain Fog
- Chronic Fatigue Syndrome
- Heart Failure
- Natto (Food Source)