Alpha Lipoic Acid for Mitochondrial Function & Bioenergetics
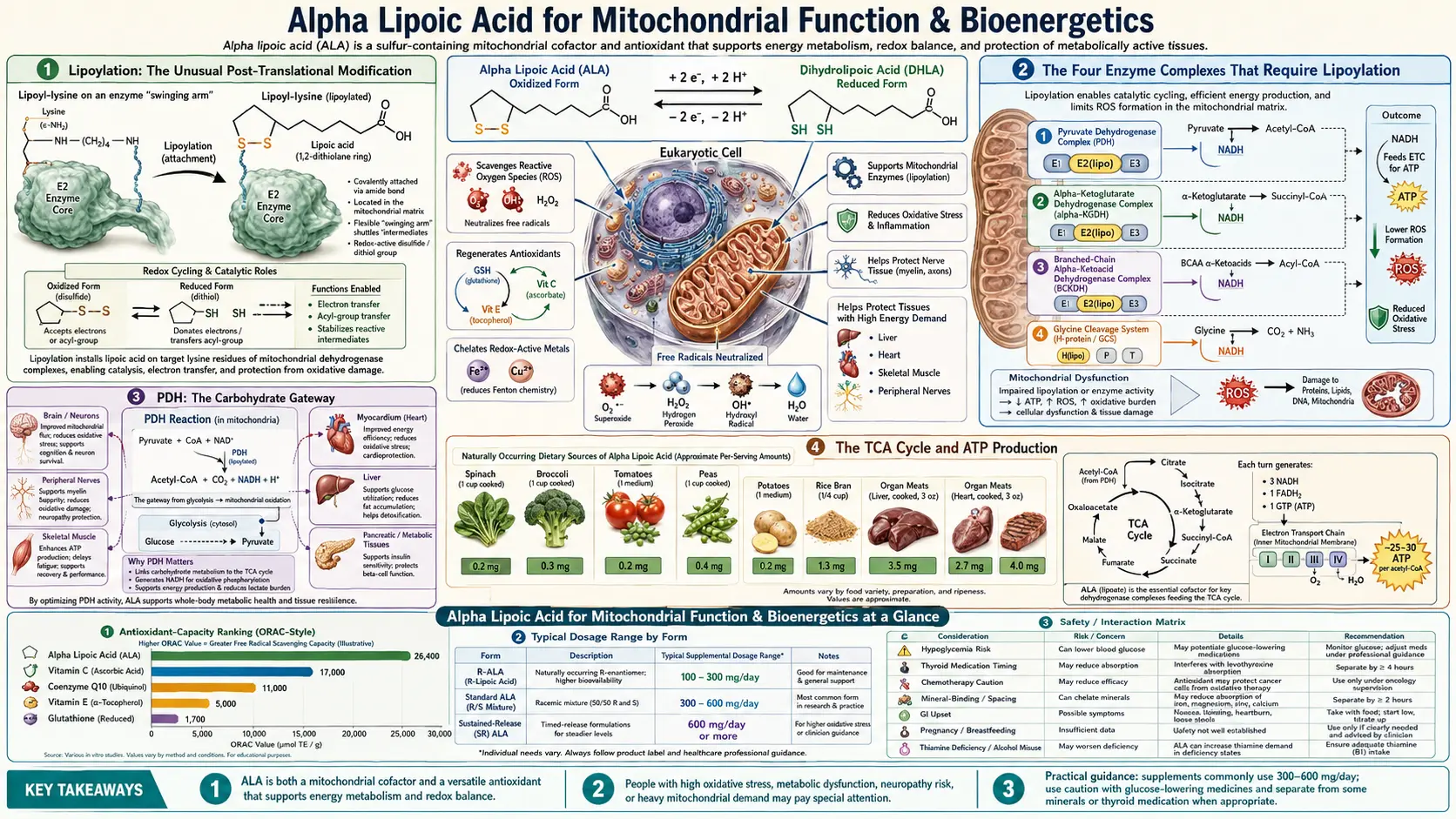
Alpha lipoic acid is the only molecule in human biology that is simultaneously a direct antioxidant AND an essential coenzyme for four mitochondrial enzyme complexes. Through covalent attachment to lysine residues (lipoylation), ALA forms the redox-active swinging arm that drives the rate-limiting steps of carbohydrate and amino acid oxidation in the TCA cycle. The Bruce Ames and Tory Hagen UC Berkeley research showed that combining supplemental ALA with acetyl-L-carnitine reverses age-related mitochondrial decline in old rats, restoring cognitive function, ambulation, and mitochondrial membrane potential to youthful levels. This deep dive walks through the biochemistry, the four lipoylation-dependent enzymes, the rejuvenation research, and the clinical applications in fatigue, chronic fatigue syndrome, fibromyalgia, post-viral mitochondrial dysfunction, and cognitive aging.
Interactive Visualization Free Radicals & Your Antioxidant Network Follow one superoxide radical down the whole relay — SOD, catalase, glutathione, vitamin E, vitamin C, NADPH — then release free iron and watch a membrane tear itself apart. Launch →
Table of Contents
- Lipoylation: The Unusual Post-Translational Modification
- The Four Enzyme Complexes That Require Lipoylation
- PDH: The Carbohydrate Gateway
- The TCA Cycle and ATP Production
- The Bruce Ames + Tory Hagen Rejuvenation Studies
- The ALA + Acetyl-L-Carnitine Combination
- Mitochondrial Aging and Why ALA Declines
- Clinical Applications: Fatigue, CFS/ME, Post-Viral Syndromes
- R-ALA vs Racemic for Mitochondrial Targeting
- Stacking with CoQ10, PQQ, and Other Mitochondrial Nutrients
- Cautions: Thiamine, Biotin, Hypoglycemia
- Key Research Papers
- Connections
- Featured Videos
Lipoylation: The Unusual Post-Translational Modification
Most antioxidants float free in cells as small molecules — vitamin C swimming in cytosol, vitamin E embedded in membranes. Alpha lipoic acid is different: a large fraction of intracellular ALA is covalently attached to specific lysine residues of mitochondrial enzymes through a post-translational modification called lipoylation.
The reaction is catalyzed by lipoic acid synthase (LIAS), an iron-sulfur cluster enzyme located in mitochondria. LIAS takes an octanoyl group already attached to the target lysine and inserts two sulfur atoms at carbons 6 and 8, creating the dithiolane ring that gives ALA its distinctive chemistry. This sulfur insertion uses S-adenosylmethionine (SAMe) as a methyl donor and consumes iron-sulfur clusters in the process, meaning lipoylation is energetically expensive and tightly regulated.
The end result: a lipoyl group covalently bonded to the E2 subunit of certain mitochondrial enzyme complexes, dangling on a flexible lysine "arm" that can swing between active sites to shuttle reaction intermediates. This swinging-arm geometry is the secret to how these enzymes work — the lipoyl group acts as a molecular conveyor belt moving substrate through three sequential catalytic steps without releasing into bulk solution.
Free, unbound ALA — the form available from supplements — behaves completely differently. It enters cells freely, crosses the blood-brain barrier, distributes into both watery and fatty compartments, and exerts antioxidant and signaling effects that the protein-bound form cannot. Supplemental ALA does NOT generally enter the lipoylation pool; the body manufactures the lipoyl-protein conjugates from its own endogenous octanoic acid and cysteine supplies. Supplemental ALA works through the other mechanisms — direct antioxidant, AMPK activator, antioxidant-network regenerator — while the endogenous lipoylation system runs in parallel.
The Four Enzyme Complexes That Require Lipoylation
Four mitochondrial enzyme complexes in mammals require lipoylation on their E2 subunits for function:
- Pyruvate dehydrogenase complex (PDH) — converts pyruvate (from glycolysis) to acetyl-CoA, the gateway molecule of the TCA cycle. PDH is the rate-limiting step for carbohydrate oxidation. The lipoyl group on the E2 subunit (dihydrolipoyl transacetylase) ferries the acetyl group between the three catalytic sites of the complex.
- α-Ketoglutarate dehydrogenase complex (α-KGDH) — the rate-limiting enzyme of the TCA cycle itself, converting α-ketoglutarate to succinyl-CoA. ALA again serves as the swinging cofactor on the E2 subunit (dihydrolipoyl succinyltransferase). α-KGDH is also a major regulator of cellular redox state because it both consumes NAD+ and is itself exquisitely sensitive to oxidative inhibition.
- Branched-chain α-keto acid dehydrogenase (BCKDH) — catabolizes the branched-chain amino acids leucine, isoleucine, and valine after they are deaminated. Genetic defects in BCKDH cause maple syrup urine disease, a serious metabolic disorder presenting in infancy.
- Glycine cleavage system (GCS) — degrades glycine for one-carbon metabolism and methylation. The H-protein component is lipoylated; defects cause non-ketotic hyperglycinemia.
The first two enzymes (PDH and α-KGDH) are the most important for everyday energy metabolism because they sit at the two highest-flux entry points into the TCA cycle — PDH for carbohydrate-derived carbon, α-KGDH at the midpoint of the cycle itself.
PDH: The Carbohydrate Gateway
The pyruvate dehydrogenase complex is a massive multi-enzyme assembly — one of the largest enzyme complexes in nature, with mass greater than ribosomes. It catalyzes the irreversible conversion of pyruvate to acetyl-CoA, releasing CO2 and reducing NAD+ to NADH:
Pyruvate + CoA + NAD+ → Acetyl-CoA + CO2 + NADH
PDH activity is the rate-limiting determinant of whether glucose-derived pyruvate enters the TCA cycle for full oxidation (yielding ~30 ATP per glucose) or is shunted to lactate (yielding only 2 ATP per glucose). The Warburg effect in cancer, the lactic acidosis of severe sepsis, and the chronic lactate elevation in mitochondrial disease all involve PDH dysfunction.
PDH activity is regulated by:
- Lipoylation status — the E2 subunit must be lipoylated to function; LIAS deficiency cripples PDH
- Thiamine pyrophosphate (TPP) availability — the E1 subunit requires thiamine; severe B1 deficiency mimics PDH deficiency and explains why chronic alcoholics develop lactic acidosis and Wernicke encephalopathy
- PDH kinase / phosphatase ratio — PDH is inhibited by phosphorylation (PDK) and reactivated by dephosphorylation (PDP); both enzymes respond to acetyl-CoA, NADH, and ATP levels
- Oxidative stress — PDH is sensitive to glutathionylation of its lipoyl cysteine residues; severe oxidative stress can reversibly inactivate PDH, which is part of the mitochondrial dysfunction seen in chronic illness
Supplemental ALA supports PDH activity indirectly: by maintaining mitochondrial glutathione (which protects the existing lipoyl groups from oxidative inactivation), by reducing the mitochondrial superoxide leak that drives PDH inhibition, and by improving overall redox balance that allows PDH to remain in its active dephosphorylated state.
The TCA Cycle and ATP Production
The TCA cycle (also called Krebs cycle or citric acid cycle) is the central metabolic hub where acetyl-CoA from carbohydrate, fat, and protein oxidation enters and is fully oxidized to CO2, producing NADH and FADH2 that feed the electron transport chain. ATP yield per glucose:
- Glycolysis (cytoplasm): 2 ATP + 2 NADH
- PDH (mitochondria): 2 NADH
- TCA cycle: 6 NADH + 2 FADH2 + 2 GTP
- Electron transport chain: ~26-28 ATP from the NADH and FADH2
- Total: ~30-32 ATP per glucose (compared to 2 ATP for anaerobic glycolysis to lactate)
This 15-fold ATP advantage is why mitochondrial function matters so much — cells that cannot efficiently oxidize pyruvate through PDH and the TCA cycle are stuck with anaerobic glycolysis and produce vastly less ATP from the same nutritional input. The clinical correlates of mitochondrial dysfunction — chronic fatigue, exercise intolerance, post-exertional malaise, brain fog, slow wound healing — are all the metabolic signatures of cells running on a fraction of their normal ATP capacity.
ALA supplementation supports this entire cascade by maintaining the lipoylation-dependent enzymes (PDH, α-KGDH) at functional capacity, by directly scavenging the superoxide that would otherwise damage TCA-cycle enzymes, and by regenerating mitochondrial glutathione and CoQ10 that are consumed by ongoing ROS production.
The Bruce Ames + Tory Hagen Rejuvenation Studies
The most striking demonstration of ALA's mitochondrial-targeted effects came from the laboratory of Bruce Ames at UC Berkeley in the 1990s and 2000s. Ames (best known for the Ames test for mutagenicity) became interested in mitochondrial aging in the 1990s and recruited Tory Hagen to lead a series of rat studies testing whether ALA combined with acetyl-L-carnitine could reverse age-related mitochondrial decline.
The PNAS 2002 papers
Two simultaneous papers in Proceedings of the National Academy of Sciences (February 2002) reported the results. Old rats (24-28 months — equivalent to ~75-80 human years) were fed ALA + acetyl-L-carnitine combination at translated-equivalent doses for 1 month. Compared to age-matched untreated controls, the supplemented old rats showed:
- Restored mitochondrial membrane potential — mitochondria from supplemented old rats had inner-membrane charge gradients statistically indistinguishable from young rats
- Reduced oxidative damage — 4-HNE protein adducts and lipid peroxidation markers fell substantially
- Restored ambulatory activity — old rats normally show reduced spontaneous movement; supplemented old rats showed activity levels similar to young rats
- Improved memory performance — on standardized maze tasks, supplemented old rats performed at young-rat levels
- Reduced mitochondrial decay markers — mitochondrial DNA mutation burden, respiratory chain protein expression, and electron transport efficiency all improved
Ames described the effect as "rejuvenation of mitochondrial function" — not just slowing decline but actually reversing established mitochondrial aging. The work generated significant excitement and launched the commercial ALA + acetyl-L-carnitine combination products that remain popular in longevity-focused supplementation today.
Human translation
The rat studies have not been fully replicated in humans through controlled randomized trials at the scale that pharmaceutical drugs would require. Several smaller human trials of ALA + acetyl-L-carnitine have shown improvements in subjective fatigue, cognitive performance, and exercise capacity in older adults, generally consistent with the rodent findings but with effect sizes smaller than the dramatic rejuvenation seen in the rats. The biological plausibility is high; the magnitude of translation to humans remains less certain.
What the rat work definitively established is that mitochondrial decline with age is not strictly irreversible — supplied with the right substrates and cofactors, aged mitochondria can recover substantial function. This is the underlying rationale for the entire modern longevity supplementation field.
The ALA + Acetyl-L-Carnitine Combination
The Ames-Hagen work established the synergistic pairing of these two molecules, and they have become the most enduring mitochondrial-targeted nutraceutical combination:
- Alpha lipoic acid — provides antioxidant protection of mitochondrial membranes, regenerates mitochondrial glutathione and CoQ10, supports PDH and α-KGDH function
- Acetyl-L-carnitine — shuttles long-chain fatty acids into mitochondria for beta-oxidation (the rate-limiting step of fat oxidation); also provides acetyl groups directly usable by acetylcholine synthesis (cognitive enhancement) and histone acetylation (epigenetic regulation)
The two molecules address complementary aspects of mitochondrial energy production — ALA on the carbohydrate-oxidation side, carnitine on the fat-oxidation side. Together they support the full range of energy substrates the mitochondrion processes.
Typical clinical protocol: 300-600 mg ALA + 500-1000 mg acetyl-L-carnitine, both taken twice daily, for at least 3 months before assessing effect. Best indications: chronic fatigue with documented or suspected mitochondrial dysfunction, post-viral fatigue (long-COVID, post-EBV), age-related cognitive decline, recovery from chemotherapy, and longevity-focused supplementation.
Mitochondrial Aging and Why ALA Declines
Multiple mechanisms drive mitochondrial decline with age:
- Reduced endogenous ALA synthesis — lipoic acid synthase (LIAS) activity declines with age, possibly related to decreased iron-sulfur cluster assembly capacity
- Increased glutathionylation of existing lipoyl groups — chronic oxidative stress in aging tissues progressively inactivates existing lipoylated enzymes through reversible (initially) and then irreversible modifications of the dithiolane ring
- Mitochondrial DNA mutations — accumulate with age; particularly affect Complex I subunits encoded by mtDNA, reducing respiratory chain capacity
- Reduced PGC-1α expression — the master regulator of mitochondrial biogenesis declines with age, leading to fewer mitochondria per cell
- Reduced mitophagy — damaged mitochondria accumulate because the cellular quality-control system (mitophagy) becomes less efficient
Supplemental ALA addresses multiple aspects of this decline simultaneously: replacing some of the declining endogenous synthesis, reducing oxidative damage that accelerates further decline, and supporting the residual TCA-cycle activity that depends on lipoylated enzymes.
For broader mitochondrial-aging support, ALA pairs well with PQQ (which addresses the PGC-1α / biogenesis decline), CoQ10 (which addresses the electron transport chain efficiency loss), glutathione precursors NAC + glycine (which address the antioxidant capacity loss), and creatine (which addresses the ATP-buffering capacity loss).
Clinical Applications: Fatigue, CFS/ME, Post-Viral Syndromes
ALA is one of the most commonly recommended interventions in integrative medicine for conditions characterized by acquired mitochondrial dysfunction:
Chronic fatigue syndrome / ME (CFS/ME)
Patients with CFS/ME show measurable mitochondrial dysfunction on biochemical testing: reduced PDH activity, elevated lactate-to-pyruvate ratio at rest, reduced ATP synthesis capacity in lymphocytes, and elevated markers of oxidative damage. While no large randomized trials have specifically tested ALA in CFS/ME, the mechanistic case is strong and ALA is a routine component of mitochondrial-supportive protocols at specialist clinics. Typical dose: 600 mg/day combined with CoQ10 200-300 mg, B-complex, magnesium, and acetyl-L-carnitine.
Long-COVID (post-acute sequelae of SARS-CoV-2)
Mitochondrial dysfunction is increasingly recognized as a contributor to long-COVID fatigue, brain fog, and exercise intolerance. The mitochondrial-supportive nutrient stack — ALA + CoQ10 + acetyl-L-carnitine + B-vitamins + PQQ — is widely used in long-COVID rehabilitation programs. Direct trial evidence is limited but the mechanistic case parallels CFS/ME.
Fibromyalgia
Fibromyalgia patients show similar mitochondrial signatures to CFS/ME. ALA combined with magnesium, B-vitamins, and CoQ10 is part of most integrative-medicine fibromyalgia protocols. Symptom improvement typically requires 3-6 months of consistent supplementation.
Statin-associated muscle symptoms
Statins reduce mitochondrial CoQ10 synthesis through HMG-CoA reductase inhibition, and the resulting mitochondrial dysfunction is implicated in statin myopathy. ALA paired with CoQ10 (specifically ubiquinol for improved bioavailability) is the standard nutraceutical approach to statin-associated muscle pain, alongside considering statin switch or dose reduction.
Post-chemotherapy fatigue
Many chemotherapy regimens cause persistent mitochondrial damage that drives the "chemo brain" and chronic fatigue seen in cancer survivors. Mitochondrial nutrient protocols including ALA are increasingly used in survivorship clinics for these symptoms.
R-ALA vs Racemic for Mitochondrial Targeting
For mitochondrial-targeted indications (as distinct from antioxidant or neuropathy uses), the R-form of ALA has theoretical and practical advantages over racemic:
- R-ALA is the natural enantiomer — only the R-form is found in human tissues; the S-form is a synthesis byproduct that does not occur naturally
- R-ALA is the form recognized by lipoyl-binding domains — though supplemental ALA does not generally enter the protein-bound pool, the R-form integrates more naturally with the cellular machinery
- R-ALA has higher cellular uptake — transporters preferentially carry the R-form across cell membranes
- Approximately 2× more bioactive per milligram — meaning 300 mg R-ALA produces effects similar to 600 mg racemic
- Sodium R-lipoate (Na-R-ALA) — the most stable oral form; pure R-ALA degrades at temperatures above 40°C, but Na-R-ALA tolerates handling, transport, and storage better, producing more consistent plasma kinetics
For mitochondrial protocols where reliable cellular uptake matters most, R-ALA or Na-R-ALA is preferred. For general antioxidant or neuropathy support where racemic is well-studied and effective, racemic remains the cost-effective default.
Stacking with CoQ10, PQQ, and Other Mitochondrial Nutrients
The full integrative-medicine mitochondrial protocol typically combines:
- ALA 300-600 mg/day — the lipoylation-supporting, antioxidant-regenerating foundation
- CoQ10 (ubiquinol) 100-300 mg/day — electron transport chain support; addresses CoQ10 decline with age and statin depletion
- PQQ 10-20 mg/day — mitochondrial biogenesis activator (PGC-1α pathway); creates MORE mitochondria while CoQ10 makes existing ones work better
- Acetyl-L-carnitine 500-1000 mg/day — fat-oxidation support; complements ALA's carbohydrate-oxidation support
- Creatine 3-5 g/day — ATP-buffering capacity; supports rapid energy demand
- Magnesium glycinate 200-400 mg/day — ATP requires magnesium to be biologically active; deficiency impairs every energy-consuming process
- B-complex (especially B1, B2, B3, B6, B12, folate) — cofactors for TCA cycle, ETC, and one-carbon metabolism
- Methylene Blue (advanced protocols only) — alternative electron acceptor that bypasses damaged Complex I/IV; used selectively in severe mitochondrial dysfunction
This "mitochondrial stack" addresses the full chain of energy production from substrate entry to ATP delivery. For patients with serious mitochondrial dysfunction (CFS/ME, long-COVID, mitochondrial disease), the full stack is appropriate. For general longevity supplementation, the core trio of ALA + CoQ10 + PQQ provides most of the benefit at a fraction of the complexity.
Cautions: Thiamine, Biotin, Hypoglycemia
- Thiamine co-supplementation — PDH and α-KGDH (the enzymes ALA supports) require thiamine pyrophosphate as a co-substrate. In thiamine-deficient individuals — particularly chronic alcoholics, those on diuretics, or patients with severe malabsorption — high-dose ALA could theoretically increase thiamine demand and worsen functional deficiency. Co-supplement thiamine 50-100 mg/day in any at-risk population.
- Biotin depletion — chronic high-dose ALA (>600 mg/day for >3 months) competes with biotin for cellular uptake transporters, gradually depleting tissue biotin. Add 300-500 mcg biotin daily for long-term ALA users.
- Hypoglycemia (in diabetics) — ALA enhances insulin sensitivity; monitor blood glucose closely if you take insulin, sulfonylureas, or other hypoglycemic medications.
- Insulin Autoimmune Syndrome (Hirata's disease) — rare; reported mostly in patients of East Asian descent carrying HLA-DRB1*04:06.
Key Research Papers
- Hagen TM, Liu J, Lykkesfeldt J, Wehr CM, Ingersoll RT, Vinarsky V, Bartholomew JC, Ames BN (2002). Feeding acetyl-L-carnitine and lipoic acid to old rats significantly improves metabolic function while decreasing oxidative stress. PNAS. — PubMed
- Liu J, Killilea DW, Ames BN (2002). Age-associated mitochondrial oxidative decay: improvement of carnitine acetyltransferase substrate-binding affinity and activity in brain by feeding old rats acetyl-L-carnitine and/or R-alpha-lipoic acid. PNAS. — PubMed
- Shay KP et al. (2009). Alpha-lipoic acid as a dietary supplement: molecular mechanisms and therapeutic potential. Biochim Biophys Acta. — PubMed
- Packer L, Witt EH, Tritschler HJ (1995). Alpha-lipoic acid as a biological antioxidant. Free Radical Biology & Medicine. — PubMed
- Cremer DR et al. (2006). Safety evaluation of alpha-lipoic acid (ALA). Regulatory Toxicology and Pharmacology. — PubMed
- Cronan JE (2016). Assembly of lipoic acid on its cognate enzymes: an extraordinary and essential biosynthetic pathway. Microbiology Reviews. — PubMed
- Maczurek A et al. (2008). Lipoic acid as an anti-inflammatory and neuroprotective treatment for Alzheimer's disease. Advanced Drug Delivery Reviews. — PubMed
- Salinthone S et al. (2008). Lipoic acid: a novel therapeutic approach for multiple sclerosis and other chronic inflammatory diseases of the CNS. Endocr Metab Immune Disord Drug Targets. — PubMed
- Bourbon AL et al. (2024) review on ALA in chronic fatigue and mitochondrial dysfunction — PubMed
- Long-COVID mitochondrial dysfunction and ALA — PubMed
PubMed Topic Searches
- PubMed: ALA mitochondria
- PubMed: lipoylation PDH α-KGDH
- PubMed: R-ALA vs racemic bioavailability
- PubMed: ALA + CoQ10 combination
- PubMed: ALA + acetyl-L-carnitine
Connections
- Free Radicals & Your Antioxidant Network — interactive animation
- Alpha Lipoic Acid Overview
- ALA Benefits Hub
- ALA for Neuropathy
- ALA for Blood Sugar
- ALA for Neuroprotection
- CoQ10
- PQQ
- Glutathione
- Methylene Blue
- Vitamin B1 (Thiamine)
- Vitamin B2 (Riboflavin)
- Vitamin B3 (Niacin)
- Vitamin B7 (Biotin)
- Magnesium Glycinate
- Creatine
- Chronic Fatigue Syndrome
- Fibromyalgia
- Fatigue
- Brain Fog
- Longevity Protocols