NAD+ & NMN for Energy and Mitochondrial Function
NAD+ is the universal electron carrier without which the mitochondrial electron transport chain cannot operate. When NAD+ falls, ATP production falls with it, reactive oxygen species leak from the chain, and cells lose the energy they need for repair, signaling, and resilience. The most direct clinical applications of NAD+ repletion therefore involve fatigue states — chronic fatigue syndrome, long-COVID, post-viral fatigue, mitochondrial myopathies, age-related sarcopenia, and the everyday fatigue of midlife. This deep-dive walks through NAD+ as electron carrier, the NAD+/NADH ratio as a bioenergetic readout, the mitochondrial unfolded protein response, AMPK and sirtuin cross-talk, the published exercise-capacity trials, the IV NAD+ infusion clinic landscape, and how to think about NMN for fatigue clinically.
Table of Contents
- NAD+ as Universal Electron Carrier
- The NAD+/NADH Ratio as Bioenergetic Indicator
- Mitochondrial Biogenesis via PGC-1α
- The Mitochondrial Unfolded Protein Response (UPRmt)
- AMPK and Sirtuin Cross-Talk
- Exercise Capacity Trials
- Chronic Fatigue Syndrome and Mitochondrial Dysfunction
- Long COVID and Post-Viral Fatigue Protocols
- IV NAD+ Infusion Clinics
- Sarcopenia and Aging Muscle
- Cardiac Energetics
- Practical Patient Protocol
- Cautions
- Key Research Papers
- Connections
- Featured Videos
NAD+ as Universal Electron Carrier
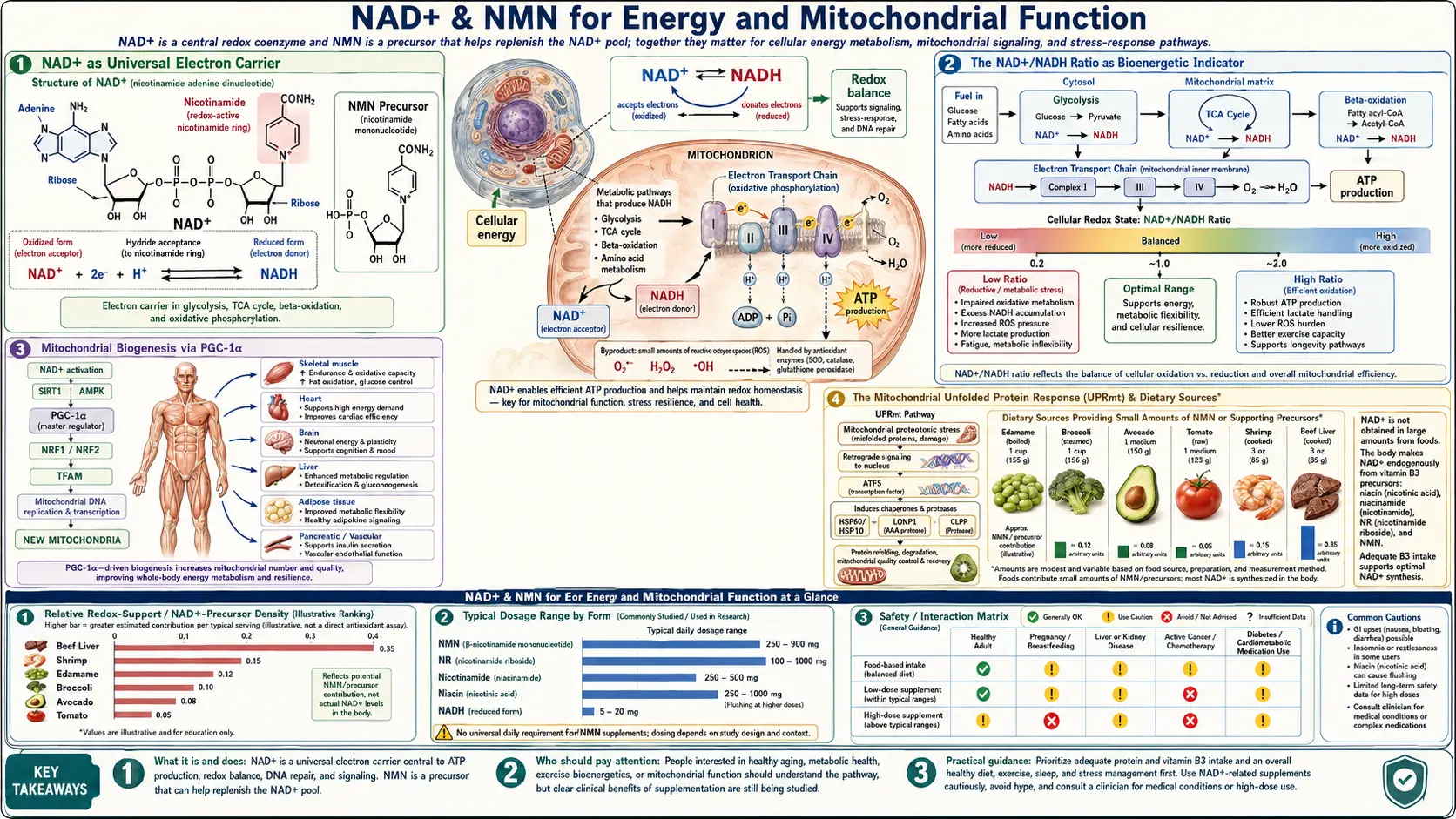
Cellular energy production reduces to a single chemical operation: capturing electrons from food molecules and using their movement down an energy gradient to drive ATP synthesis. NAD+ is the molecule that does the capturing. During glycolysis, the citric acid cycle, and fatty acid β-oxidation, NAD+ accepts electrons (and a proton) from carbon-bearing substrates to become NADH. The NADH then carries those electrons to Complex I of the mitochondrial electron transport chain, where they are passed down through Complexes I, III, and IV while pumping protons across the inner mitochondrial membrane. The proton gradient drives ATP synthase, and ATP is produced.
The arithmetic of the system explains why NAD+ is so central:
- Glycolysis (one glucose to two pyruvate) generates 2 NADH
- Pyruvate dehydrogenase (two pyruvate to two acetyl-CoA) generates 2 NADH
- Citric acid cycle (two acetyl-CoA fully oxidized) generates 6 NADH and 2 FADH2
- Each NADH that enters the electron transport chain at Complex I yields ~2.5 ATP
Without enough NAD+, glycolysis stalls (no acceptor for the GAPDH-generated electrons), pyruvate cannot be oxidized to acetyl-CoA, the citric acid cycle slows, and cellular ATP production collapses. The cell then resorts to anaerobic glycolysis (lactate production), which yields only 2 ATP per glucose instead of ~36, and acidifies the cytoplasm.
This is why NAD+ depletion is so consequential: it does not just dial down one pathway but disables the entire oxidative metabolic apparatus on which cells depend for sustained energy production.
The NAD+/NADH Ratio as Bioenergetic Indicator
The cell does not just care about the absolute amount of NAD+ — it cares about the ratio of NAD+ to NADH. This ratio is the bioenergetic readout that triggers downstream signaling. A high NAD+/NADH ratio (more oxidized form) signals fasting, exercise, caloric restriction, and metabolic stress; a low NAD+/NADH ratio (more reduced form) signals fed state, abundant fuel availability, and storage mode.
The NAD+/NADH ratio influences cellular function through:
- Sirtuin activity — sirtuins require NAD+ specifically (not NADH) as substrate. A high NAD+/NADH ratio means active sirtuins and the caloric-restriction-mimetic phenotype: autophagy, mitochondrial biogenesis, stress resistance, anti-inflammatory signaling.
- SIRT3 in mitochondria — the mitochondrial NAD+/NADH ratio drives SIRT3, which deacetylates and activates the antioxidant defenses and the fatty acid oxidation enzymes.
- Pyruvate-lactate balance — lactate dehydrogenase uses the cytoplasmic NAD+/NADH ratio to set the pyruvate-to-lactate equilibrium. Low NAD+/NADH (high NADH) drives pyruvate to lactate, contributing to lactic acidosis in mitochondrial dysfunction states.
- Gluconeogenesis — the liver's ability to make glucose from non-carbohydrate precursors during fasting depends on the cytoplasmic NAD+/NADH ratio.
NMN/NR supplementation raises the absolute NAD+ pool, which in most contexts raises the NAD+/NADH ratio as well. This is the biochemical basis for the framing of NMN as a fasting mimetic or exercise mimetic.
Mitochondrial Biogenesis via PGC-1α
Beyond serving as electron carrier, NAD+ regulates the number and quality of mitochondria in each cell through the PGC-1α (peroxisome proliferator-activated receptor gamma coactivator 1-alpha) pathway. PGC-1α is the master regulator of mitochondrial biogenesis — the cellular program for assembling new mitochondria from scratch.
The pathway works as follows:
- Caloric restriction, exercise, or other energy stress raises the cellular NAD+/NADH ratio.
- Elevated NAD+ activates SIRT1.
- Activated SIRT1 deacetylates PGC-1α, converting it from an inactive to an active state.
- Active PGC-1α drives transcription of nuclear-encoded mitochondrial genes (mitochondrial transcription factor A, respiratory chain subunits, fatty acid oxidation enzymes).
- New mitochondria are assembled, increasing cellular oxidative capacity.
This is the molecular basis of one of the foundational principles of exercise physiology: aerobic training increases mitochondrial density in skeletal muscle, which then enables greater endurance capacity. NAD+ availability is the rate-limiting step at the SIRT1 deacetylation node. NMN supplementation has been shown in mouse studies to increase mitochondrial density in skeletal muscle, particularly when combined with exercise training.
AMPK provides a parallel pathway: AMPK phosphorylates PGC-1α at sites that prime it for SIRT1-mediated deacetylation. NAD+ and AMPK therefore cooperate to drive mitochondrial biogenesis (see the AMPK section below).
The Mitochondrial Unfolded Protein Response (UPRmt)
Mitochondria have their own quality-control system that monitors the folding state of their proteins and triggers a stress response when too many are misfolded. This system, called the mitochondrial unfolded protein response (UPRmt), is the mitochondrial equivalent of the better-known endoplasmic reticulum UPR.
The UPRmt was first characterized in C. elegans, where mild activation of the response extends lifespan substantially. Subsequent work in mammals has shown that the UPRmt is regulated in part by NAD+/sirtuin signaling and that it activates a coordinated program including:
- Mitochondrial chaperone induction (mtHSP70, HSP60) to refold misfolded proteins
- Mitochondrial protease induction (LonP1, ClpXP) to degrade terminally misfolded proteins
- Mitochondrial biogenesis to replace damaged mitochondria with new ones
- Reduced cytoplasmic translation to lower the demand on mitochondrial protein import machinery
- Mitochondrial-derived peptide signaling (e.g., MOTS-c) to non-mitochondrial systems
Mouse and worm studies have shown that NAD+ precursor supplementation activates the UPRmt and produces lifespan and healthspan benefits in part through this pathway. Mitonuclear protein imbalance, induced by NAD+ depletion or directly by genetic manipulation, triggers UPRmt and extends lifespan. The UPRmt is therefore one of the proposed mechanisms by which NMN and NR produce their longevity effects in preclinical models.
AMPK and Sirtuin Cross-Talk
AMPK (AMP-activated protein kinase) and the sirtuins are the cell's two principal energy-status sensors, and they cooperate extensively. AMPK is activated when the AMP/ATP ratio rises (signaling low energy); sirtuins are activated when the NAD+/NADH ratio rises (signaling oxidative metabolic stress).
The cross-talk runs in both directions:
- AMPK phosphorylates and activates NAMPT, increasing NAD+ salvage synthesis — raising NAD+ availability for sirtuins.
- AMPK phosphorylates PGC-1α, priming it for SIRT1-mediated deacetylation.
- Sirtuins deacetylate LKB1, the upstream activator of AMPK, increasing AMPK activity.
- SIRT1 deacetylates PGC-1α that has been phosphorylated by AMPK, fully activating it for transcriptional duty.
The clinical consequence: interventions that activate either pathway tend to amplify the other. Caloric restriction, exercise, metformin (an AMPK activator), berberine (also AMPK), and NMN/NR (sirtuin substrate) all converge on the same downstream metabolic adaptation. This is why integrative practitioners sometimes combine NMN with metformin or berberine for metabolic-syndrome populations — the combinations are mechanistically synergistic.
It is also why exercise is the single most powerful endogenous activator of the entire system: a single aerobic exercise session raises AMPK, raises the NAD+/NADH ratio, activates sirtuins, and triggers PGC-1α-driven mitochondrial biogenesis. No supplement matches the effect.
Exercise Capacity Trials
The most rigorous human exercise data on NMN come from Liao et al. (2021), a randomized double-blind dose-ranging trial in 48 young to middle-aged recreational runners. Participants received placebo, 300 mg, 600 mg, or 1,200 mg NMN per day for 6 weeks alongside endurance training 5–6 days per week.
Results:
- Ventilatory threshold (the work intensity at which lactate begins to accumulate) increased significantly in the 600 mg and 1,200 mg groups but not the 300 mg group
- Oxygen uptake at submaximal intensities increased in the higher-dose groups
- The percentage of VO2max at which participants could work increased dose-dependently
- VO2max itself, peak power output, and maximal ventilation did not change significantly
- Safety: NMN up to 1,200 mg/day was well tolerated; no serious adverse events
The pattern suggests NMN preferentially benefits submaximal sustained endurance performance — the energy system most dependent on oxidative phosphorylation — rather than peak anaerobic power. This is consistent with the proposed mechanism of enhanced mitochondrial function and improved oxidative substrate handling.
The Uthever multicenter NMN trial in middle-aged and older adults also showed significant improvement on a six-minute walking endurance test. A separate February 2026 trial in healthy men found NMN reduced inflammatory markers after intense exercise, suggesting potential benefits for recovery.
For most recreational athletes, the takeaway is modest: at doses of 600 mg/day or higher, NMN may produce a measurable improvement in sustained endurance performance, particularly when combined with regular training. The effect is unlikely to be transformative, and its magnitude should not be overstated.
Chronic Fatigue Syndrome and Mitochondrial Dysfunction
Myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS) is the prototypical chronic fatigue disorder — a complex, multisystem illness characterized by post-exertional malaise, unrefreshing sleep, cognitive dysfunction, and severe functional impairment lasting more than six months. A consistent biological finding in ME/CFS is impaired mitochondrial function: reduced ATP production from peripheral blood mononuclear cells, increased lactate production with mild exertion, and abnormal cellular bioenergetic profiles on Seahorse XF analysis.
The mitochondrial dysfunction in ME/CFS has multiple proposed contributors:
- Oxidative damage to mitochondrial DNA and respiratory chain proteins from chronic inflammation
- Reduced NAD+ availability from chronic immune activation (CD38 upregulation, PARP activation from chronic DNA damage)
- Abnormal pyruvate dehydrogenase activity reducing acetyl-CoA generation
- Impaired fatty acid oxidation
- Reactivation of latent herpesviruses (EBV, HHV-6) producing chronic immune activation
The rationale for NAD+ precursors in ME/CFS is straightforward: replenish the NAD+ pool, support the impaired bioenergetic machinery, and reduce the chronic inflammation that drives ongoing NAD+ depletion. Clinical evidence is anecdotal and preliminary — no large RCT has tested NMN or NR specifically in ME/CFS — but many integrative practitioners include NMN/NR (typically with CoQ10, D-ribose, magnesium, and L-carnitine) in their ME/CFS protocols. See our Chronic Fatigue Syndrome page for the broader clinical picture.
Long COVID and Post-Viral Fatigue Protocols
The post-acute sequelae of viral infection — including the syndrome popularly known as long COVID and the broader category of post-viral fatigue syndromes following EBV, mononucleosis, Ross River virus, and others — share substantial overlap with ME/CFS in symptomatology and proposed mechanism. Mitochondrial dysfunction, persistent low-grade immune activation, and reduced NAD+ availability appear common across the post-viral fatigue spectrum.
Many integrative-medicine practitioners have used NMN, NR, and IV NAD+ infusions as part of post-viral fatigue protocols. The biological rationale is supported by:
- Documented NAD+ depletion in COVID-19 (both acute and post-acute), driven by PARP activation from viral-induced DNA damage and by CD38 upregulation from chronic immune activation
- Mitochondrial dysfunction in long-COVID patients including reduced oxidative capacity and increased lactate production
- Mechanistic case that restoring NAD+ should support recovery of mitochondrial function
Rigorous clinical trial evidence remains limited, however. Anecdotal reports of benefit are common but not equivalent to controlled data. NMN/NR for post-viral fatigue should be approached as a reasonable mechanistic intervention with modest evidence base, not as proven treatment. The basic principles of post-viral fatigue management — activity pacing, sleep optimization, gradual rather than sudden return to exercise, addressing comorbid conditions like POTS and MCAS — remain the foundation.
IV NAD+ Infusion Clinics
Intravenous NAD+ infusion has emerged as a popular wellness offering, particularly in longevity clinics, addiction treatment centers, and IV nutrient therapy practices. A typical IV NAD+ session involves 250–1,000 mg of NAD+ delivered over 2–4 hours, often as part of a multi-day protocol. Costs typically range from $500 to over $1,500 per session.
The scientific basis for IV NAD+ is contested:
Arguments for IV NAD+
- Bypasses gut absorption, gut microbial deamidation, and first-pass liver metabolism
- Achieves much higher blood NAD+ concentrations than oral precursors
- Subjective reports of improved energy, mental clarity, and reduced cravings (particularly in addiction recovery contexts)
- A 2019 pilot study in elderly adults showed IV NAD+ was well tolerated and safe
Arguments against IV NAD+
- NAD+ itself is a large charged molecule that cannot easily cross cell membranes, so intravenous NAD+ may simply be degraded extracellularly to NMN or NAM before any cellular uptake occurs
- If the relevant precursor is actually NMN or NAM, then oral NMN at a fraction of the cost would achieve similar intracellular outcomes
- The infusion experience is often unpleasant (flushing, nausea, chest tightness) when run faster than 2–4 hours
- Not FDA-approved for any indication; exists in a regulatory gray area
- Cost is substantially higher than oral supplementation for unclear additional benefit
- Limited rigorous trial data
The honest assessment: IV NAD+ produces a dramatic acute experience of higher blood NAD+ and (often) improved subjective energy, but whether this translates to meaningful long-term clinical benefit beyond what oral precursors achieve is not established. For most users, oral NMN/NR at $30–60/month appears to provide similar biochemical effects as IV NAD+ infusions at $500–1,500/session, with the caveat that direct comparison studies are lacking.
Cases where IV NAD+ may have clearer rationale:
- Severe chronic fatigue states where oral precursors have not produced response
- Addiction recovery contexts (some clinical experience suggests acute reduction in cravings)
- Acute post-viral fatigue syndromes when oral absorption may be impaired
- Patients unable to tolerate oral supplementation
Sarcopenia and Aging Muscle
Sarcopenia — the progressive loss of skeletal muscle mass, strength, and function with aging — is one of the most consequential aging-related changes, contributing to falls, fractures, disability, and mortality. Mitochondrial dysfunction is central to sarcopenia: aged skeletal muscle shows reduced mitochondrial density, impaired oxidative phosphorylation, and decreased oxidative capacity.
NAD+ repletion has been tested in aging-muscle contexts:
- Mouse studies: NMN restored mitochondrial function in aged skeletal muscle within one week of administration (Gomes et al., Cell 2013) and improved muscle function in aged mice over longer treatment courses
- Human exercise trials: NMN improved submaximal endurance performance in older recreational runners
- The Igarashi 2022 NMN trial in older men showed modest improvements in muscle function measures
- Yoshino 2021 showed improved skeletal muscle insulin signaling in prediabetic women
Resistance exercise remains the single most effective intervention for sarcopenia. NMN supplementation appears to be a reasonable adjunct to resistance training but is not a substitute for it. The combination of resistance training, adequate protein intake (1.2–1.6 g/kg/day for older adults), and NAD+ precursor supplementation is the most-defensible integrated approach for preserving muscle into late life.
Cardiac Energetics
The heart beats approximately 100,000 times per day and requires sustained, reliable ATP production to do so. Heart failure, whether with reduced ejection fraction (HFrEF) or preserved ejection fraction (HFpEF), is increasingly understood as a disease of cardiac energetics: the failing heart cannot meet its ATP demand because of impaired mitochondrial function.
Cardiac NAD+ levels decline with age and decline further in heart failure. SIRT3, the mitochondrial sirtuin, is particularly important in cardiac myocytes. Preclinical studies have shown that boosting NAD+ levels with NMN or NR improves cardiac function in mouse models of:
- Ischemic cardiomyopathy
- Diabetic cardiomyopathy
- Dilated cardiomyopathy
- Hypertrophic cardiomyopathy
- Arrhythmogenic cardiomyopathy
- Heart failure with preserved ejection fraction
Human cardiac trials are emerging. The Martens 2018 NR trial showed modest reductions in systolic blood pressure and aortic stiffness in middle-aged adults. A Harvard-affiliated NMN trial reported reduction in diastolic blood pressure and body weight in overweight middle-aged adults. Larger and disease-specific cardiac trials are in progress.
For patients with cardiovascular disease, NMN/NR should be considered an adjunct to standard cardiac care (statins, ACE inhibitors, beta-blockers, SGLT2 inhibitors, diet, exercise), not a substitute. The mechanistic case is strong; the clinical-trial case is developing.
Practical Patient Protocol
For general energy support / midlife fatigue
- NMN 250–500 mg/day or NR 300–500 mg/day
- Morning, empty stomach
- Expect 4–12 weeks before assessing response; subjective energy is the primary endpoint
- Co-supplement TMG 500 mg/day
For chronic fatigue / post-viral fatigue
- NMN 500–1,000 mg/day may be needed at the higher end
- Combine with CoQ10 200–400 mg/day (ubiquinol form preferred over ubiquinone)
- D-ribose 5 g three times daily
- L-carnitine 1–2 g/day (or acetyl-L-carnitine 1–2 g/day for CNS-prominent fatigue)
- Magnesium glycinate 200–400 mg/day
- Address comorbid conditions (POTS, MCAS, SIBO, hypothyroidism, low iron) that drive ongoing fatigue
For exercise capacity / athletic performance
- NMN 600–1,200 mg/day per the Liao 2021 dose-response data
- Take 30–60 minutes before training on training days
- Combine with regular endurance training (the supplement does not substitute for the training)
For cardiac support
- NMN 500–1,000 mg/day or NR 500–1,000 mg/day
- Combine with CoQ10 (especially if on a statin)
- Maintain standard cardiac care
Cautions
- Hypoglycemia in insulin-treated diabetics — NMN can enhance insulin sensitivity and lower blood glucose. Monitor closely in the first 2–4 weeks of supplementation if on insulin or sulfonylureas; expect to potentially reduce diabetes medication doses.
- Sleep disturbance from late-day dosing — the energy-promoting effects of NMN can interfere with sleep onset if taken in the afternoon or evening. Take in the morning.
- Methylation depletion at chronic high dose — add TMG 500–1,000 mg/day if taking NMN/NR above 500 mg/day chronically. Monitor homocysteine.
- IV NAD+ infusions can be unpleasant — chest tightness, abdominal discomfort, flushing, and nausea are common during infusion. These usually resolve when the infusion rate is slowed. Discuss with your provider.
- Heart failure patients should not self-medicate — NMN/NR is reasonable adjunct in cardiac disease, but coordinate with your cardiologist; do not stop standard heart failure medications.
- Theoretical cancer concern — see Longevity & Sirtuins for the discussion.
- Long-term safety unknown — no NMN/NR trial has run longer than 24 weeks; multi-year effects are uncharacterized.
Key Research Papers
- Liao, B., et al. (2021). Nicotinamide Mononucleotide Supplementation Enhances Aerobic Capacity in Amateur Runners: A Randomized, Double-Blind Study. Journal of the International Society of Sports Nutrition 18(1), 54. — DOI
- Gomes, A.P., et al. (2013). Declining NAD+ Induces a Pseudohypoxic State Disrupting Nuclear-Mitochondrial Communication During Aging. Cell 155(7), 1624–1638. — PubMed
- Mills, K.F., et al. (2016). Long-Term Administration of Nicotinamide Mononucleotide Mitigates Age-Associated Physiological Decline in Mice. Cell Metabolism 24(6), 795–806. — DOI
- Yoshino, M., et al. (2021). Nicotinamide Mononucleotide Increases Muscle Insulin Sensitivity in Prediabetic Women. Science 372(6547), 1224–1229. — DOI
- Martens, C.R., et al. (2018). Chronic Nicotinamide Riboside Supplementation Is Well-Tolerated and Elevates NAD+ in Healthy Middle-Aged and Older Adults. Nature Communications 9(1), 1286. — DOI
- Igarashi, M., et al. (2022). Chronic Nicotinamide Mononucleotide Supplementation Elevates Blood NAD Levels and Alters Muscle Function in Healthy Older Men. NPJ Aging 8, 5. — PubMed
- Camacho-Pereira, J., et al. (2016). CD38 Dictates Age-Related NAD Decline and Mitochondrial Dysfunction Through a SIRT3-Dependent Mechanism. Cell Metabolism 23(6), 1127–1139. — DOI
- Houtkooper, R.H., et al. (2013). Mitonuclear Protein Imbalance as a Conserved Longevity Mechanism. Nature 497(7450), 451–457. — PubMed
- Canto, C., et al. (2009). AMPK Regulates Energy Expenditure by Modulating NAD+ Metabolism and SIRT1 Activity. Nature 458(7241), 1056–1060. — PubMed
- Tomas-Loba, A., et al. (2008). Telomerase Reverse Transcriptase Delays Aging in Cancer-Resistant Mice. Cell. — context for sirtuin-telomere interaction — PubMed
- Grant, R., et al. (2019). A Pilot Study Investigating Changes in the Human Plasma and Urine NAD+ Metabolome During a 6-Hour Intravenous Infusion of NAD+. Frontiers in Aging Neuroscience. — PubMed
- Bonkowski, M.S., Sinclair, D.A. (2016). Slowing Ageing by Design: The Rise of NAD+ and Sirtuin-Activating Compounds. Nature Reviews Molecular Cell Biology 17(11), 679–690. — DOI
PubMed Topic Searches
- PubMed: NMN exercise capacity
- PubMed: NAD+ ME/CFS
- PubMed: NAD+ long-COVID post-viral
- PubMed: IV NAD+ clinical
- PubMed: SIRT3 cardiac mitochondria
- PubMed: AMPK-SIRT1 mitochondrial biogenesis
Connections
- NAD+ & NMN Overview
- NAD+ Benefits Hub
- NAD+ for Longevity & Sirtuins
- NAD+ for Cognition
- NAD Precursors Compared
- CoQ10
- Alpha Lipoic Acid for Mitochondria
- All Antioxidants
- Chronic Fatigue Syndrome
- Cardiovascular Disease
- Insulin Resistance
- Longevity Protocols
- Berberine (AMPK)
- Fasting
- Creatine
- Vitamin B3 (Niacin)
- Magnesium