Arginine for Immune Function
Arginine occupies a strategic position in immunology that no other amino acid shares. It is the obligate substrate for inducible nitric oxide synthase (iNOS, NOS2) in macrophages — the antimicrobial effector pathway by which phagocytes kill intracellular bacteria, parasites, and tumor cells. It is required for T-cell receptor zeta chain expression — without arginine, the TCR signaling complex literally cannot assemble, and T cells become functionally paralyzed. It is the substrate that myeloid-derived suppressor cells (MDSCs) in tumors deplete from the microenvironment to disable nearby T cells — an immune-evasion strategy that has emerged as one of the most important mechanisms of cancer immunosuppression and is now a target of multiple oncology drugs. And in septic patients, the depletion of plasma arginine by upregulated arginase is a recognized component of the immune dysfunction that contributes to ICU mortality. The clinical translation has produced the perioperative immunonutrition literature (consistently positive for elective surgery), a more nuanced critical care literature (positive in elective surgery, mixed in sepsis), and the rapidly evolving field of arginine-pathway oncology that targets MDSC arginase to restore anti-tumor T-cell function. This deep-dive walks through each of these mechanisms and the clinical evidence that connects them.
Table of Contents
- Arginine in T-Cell Activation and TCR Zeta Chain
- M1/M2 Macrophage Polarization
- Natural Killer Cell Function
- The MDSC Arginase Mechanism in Cancer
- Targeting MDSC Arginase Therapeutically
- Septic Patient Arginine Deficiency
- ICU Immunonutrition Trials
- Elective Surgery Immunonutrition
- HIV and Chronic Viral Infection
- Sickle Cell Disease and Vaso-Occlusive Crisis
- Asthma and the Th2/Arginase Axis
- Key Research Papers
- Connections
- Featured Videos
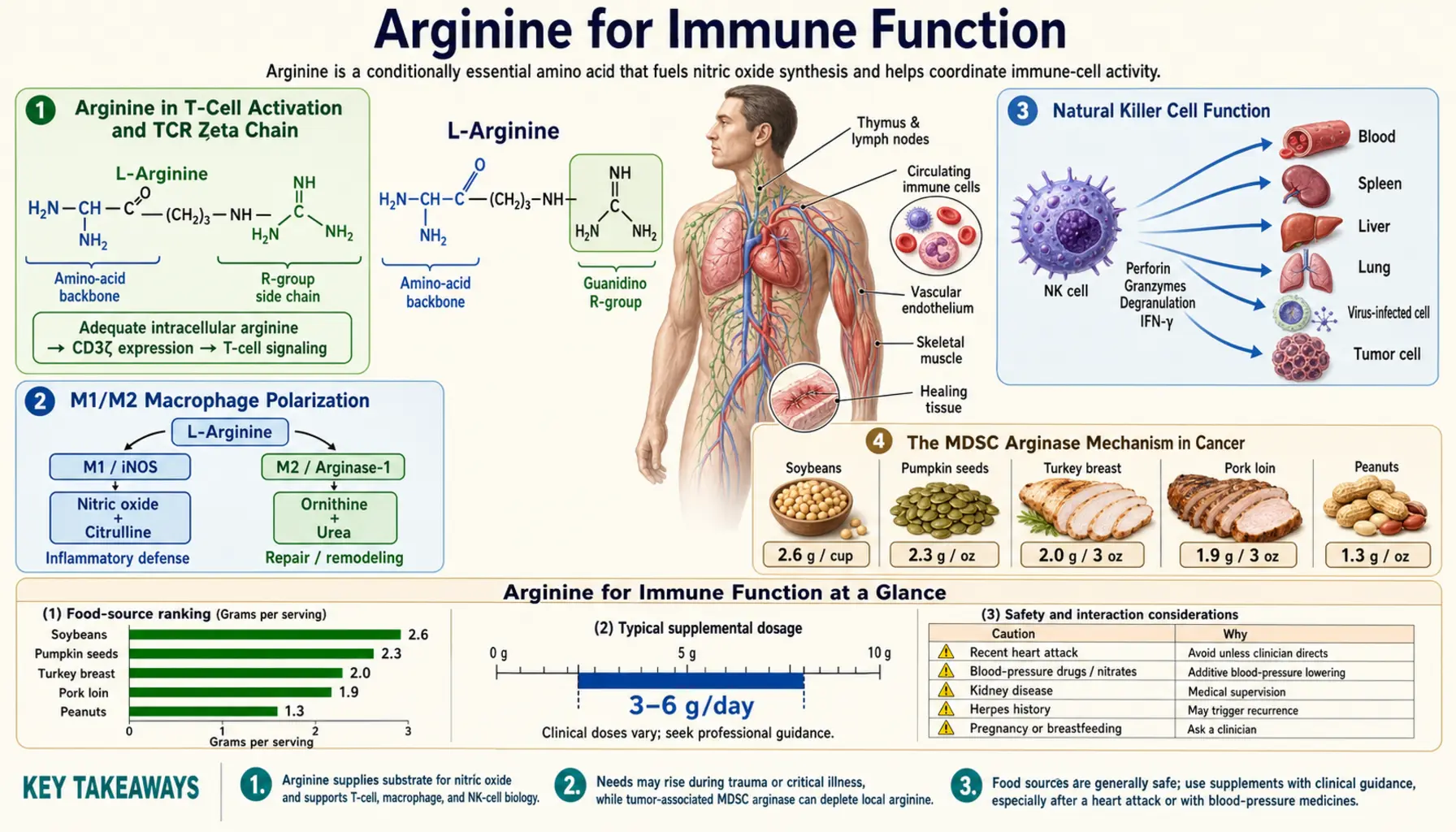
Arginine in T-Cell Activation and TCR Zeta Chain
One of the most striking and counterintuitive findings in arginine immunology is that T cells require extracellular arginine for proper function, not because they consume it for protein synthesis (they could draw on intracellular arginine for that), but because arginine specifically regulates the expression of the T-cell receptor zeta chain (CD3-zeta, CD247) — the essential signaling component of the TCR complex.
The TCR is not a single molecule but a complex of multiple subunits: the antigen-recognizing alpha-beta or gamma-delta TCR heterodimer, the CD3 complex of gamma, delta, and epsilon chains, and the homodimeric CD3-zeta chain. Of these, CD3-zeta contains three immunoreceptor tyrosine-based activation motifs (ITAMs) that are the proximal docking sites for the kinase ZAP-70 upon TCR engagement. Without CD3-zeta, antigen recognition by the TCR cannot translate into downstream signaling, calcium flux, NFAT activation, IL-2 production, or proliferation. T cells lacking CD3-zeta look intact on the surface but are functionally paralyzed.
The work of Augusto Ochoa, Paulo Rodriguez, and colleagues at LSU Health Sciences Center demonstrated that arginine deprivation downregulates CD3-zeta expression at the post-transcriptional level. T cells cultured in arginine-deficient media show normal levels of CD3-zeta mRNA but dramatically reduced CD3-zeta protein, with translation specifically blocked. The mechanism involves the GCN2 amino acid stress response kinase, which phosphorylates eIF2-alpha and selectively inhibits the translation of mRNAs containing specific upstream open reading frames — CD3-zeta mRNA happens to be one of these regulated targets.
The functional consequence is that T cells in an arginine-deficient microenvironment lose their ability to respond to antigen. They survive, they still express their TCR on the surface, but they cannot proliferate or produce cytokines in response to stimulation. This is a relatively new (post-2002) understanding that has rewritten how immunologists think about the impact of nutritional status on adaptive immunity — it's not just about generic protein-energy malnutrition; arginine specifically gates T-cell function through a defined molecular mechanism.
The implication for clinical immunology is that any situation that depletes arginine in the local T-cell microenvironment will produce functional immunodeficiency, even if circulating lymphocyte counts and surface phenotypes look normal. The tumor microenvironment is the most-studied such situation (discussed below), but the same mechanism contributes to immune dysfunction in sepsis, post-traumatic immunosuppression, severe burns, and chronic inflammation.
M1/M2 Macrophage Polarization
Macrophages are not a single cell type but a continuum of activation states. The two extreme phenotypes are:
- M1 (classically activated, pro-inflammatory) — induced by Th1 cytokines (interferon-gamma, TNF-alpha, LPS). High iNOS expression, high NO production, high pro-inflammatory cytokine output (IL-1, IL-6, IL-12, TNF), antimicrobial and anti-tumor effector function. Uses arginine via iNOS to produce NO.
- M2 (alternatively activated, pro-resolving) — induced by Th2 cytokines (IL-4, IL-13). High arginase expression, high ornithine/polyamine/proline production, tissue repair and angiogenesis functions, often immunosuppressive. Uses arginine via arginase to produce ornithine.
The M1 vs M2 dichotomy is the centerpiece of arginine immunology. Both phenotypes consume arginine, but they route it down opposite metabolic paths to opposite functional ends. The M1 macrophage uses arginine to kill (NO is cytotoxic); the M2 macrophage uses arginine to build (proline for collagen, polyamines for cell division). The wound healing literature exploits the M1-to-M2 transition that occurs over days 1–7 of a healing wound — sterilize first (M1, NO), rebuild second (M2, proline).
This dichotomy also explains why arginine has paradoxical effects in different inflammatory states:
- In an acute infection with M1-dominant macrophage response, supplementing arginine enhances iNOS-driven antimicrobial NO production. Beneficial.
- In a chronic Th2-driven inflammation (asthma, parasitic infection) with M2-dominant macrophage response, supplementing arginine may feed pathological arginase-driven tissue remodeling and reduce local arginine available to NOS. Potentially harmful or beneficial depending on context.
- In a tumor microenvironment with MDSC-dominant arginase activity, the depletion of arginine paralyzes anti-tumor T cells; supplementing systemic arginine is unlikely to overcome the local depletion, and the more rational therapeutic target is the MDSC arginase itself.
The clinical takeaway: arginine immunology is context-dependent. "Boost the immune system with arginine" is not the right mental model. The right model is: identify which immune cell population is rate-limited by arginine availability in a given clinical state, and target the supplementation (or arginase inhibition) accordingly.
Natural Killer Cell Function
Natural killer (NK) cells are the principal innate-immune defense against virally infected cells and tumor cells. They detect "missing self" (cells that have downregulated MHC class I, often as a viral or tumor immune-evasion strategy) and kill targets via perforin-granzyme release and Fas-FasL interactions. NK cells are an important component of cancer immunosurveillance and contribute to viral immunity.
Arginine is required for NK cell cytotoxicity. NK cells express CD3-zeta as part of activating receptor complexes (NKp30, NKp46, CD16), and the arginine-CD3-zeta mechanism that operates in T cells also gates NK cell function. Arginine-deprived NK cells show reduced cytotoxic activity against tumor targets in vitro, and arginine supplementation restores function.
The clinical translation has been studied in cancer patients (where NK cell function is often impaired by tumor-derived arginine depletion via MDSCs), in elderly patients (where NK function declines with immunosenescence), and in chronic viral infection. Arginine supplementation as part of immunonutrition has been associated with partial restoration of NK cytotoxic function in some of these populations, though the effect sizes are modest and the clinical correlates (reduced infection rate, improved tumor response) are not consistently demonstrated.
The MDSC Arginase Mechanism in Cancer
Myeloid-derived suppressor cells (MDSCs) are a heterogeneous population of immature myeloid cells (granulocytic and monocytic subtypes) that expand dramatically in cancer patients and accumulate in tumors, secondary lymphoid organs, and the circulation. MDSCs are one of the most important mechanisms of tumor-driven immune suppression and are an active target of anti-cancer drug development.
The two principal mechanisms by which MDSCs suppress anti-tumor immunity are:
- Arginase 1 secretion — MDSCs express high levels of arginase 1, which is released into the tumor microenvironment and rapidly depletes local arginine. The depletion downregulates CD3-zeta on infiltrating T cells (the mechanism discussed above), functionally paralyzing them. Anti-tumor T cell responses cannot proceed.
- iNOS expression — MDSCs also express iNOS, producing NO that inhibits T cell proliferation through several mechanisms including nitration of signaling proteins, suppression of IL-2 receptor signaling, and induction of T-cell apoptosis.
The combination produces a profound local immunosuppression around the tumor: arginine is depleted by arginase, what little NO is produced via iNOS is itself immunosuppressive at high local concentrations, and the T cells that should be attacking the tumor are rendered functionally inert. This is one of the leading mechanisms by which tumors evade immune surveillance despite presenting numerous tumor-associated antigens that should be immunogenic.
The MDSC arginase mechanism is now recognized as a major driver of resistance to immune checkpoint inhibitor therapy (pembrolizumab, nivolumab, ipilimumab and similar drugs). Patients whose tumors have high MDSC infiltration tend to be resistant to checkpoint blockade, presumably because even when the checkpoint inhibition releases the brakes on T cells, the arginine-depleted microenvironment prevents the released T cells from mounting an effective response.
Targeting MDSC Arginase Therapeutically
The recognition of MDSC arginase as a major mechanism of tumor immunosuppression has launched an active drug development program. Several approaches are in clinical trials:
- Arginase inhibitors — small molecules such as CB-1158 (numidargistat, INCB001158) that competitively inhibit arginase activity. CB-1158 has been studied in combination with checkpoint inhibitors in melanoma, colorectal cancer, and other solid tumors. The hypothesis is that arginase inhibition restores arginine concentration in the tumor microenvironment, restores T-cell function, and amplifies checkpoint blockade efficacy.
- Arginine supplementation — the simpler approach of raising systemic arginine concentration to overcome local depletion. The biochemistry suggests this is unlikely to work in established tumors with robust MDSC infiltration (the local arginase capacity exceeds any reasonable supplementation strategy), but may have a role in preventing MDSC-driven suppression in earlier stages of tumor development or in supportive care.
- MDSC depletion strategies — targeting the MDSCs themselves with antibodies, kinase inhibitors, or differentiation agents (all-trans retinoic acid has been used to drive MDSC differentiation toward less suppressive phenotypes).
- iNOS inhibitors — targeting the parallel iNOS-driven immunosuppression. The challenge is achieving tumor-selective iNOS inhibition without compromising NO signaling in the cardiovascular system and other tissues where it is beneficial.
The clinical impact of arginase-targeting drugs in oncology is still emerging. Early-phase trials have shown that CB-1158 can be combined safely with pembrolizumab and produces measurable changes in tumor T-cell infiltration. Definitive efficacy data in larger trials is pending. Regardless of the specific drugs that ultimately succeed in this space, the arginine-immunology paradigm has fundamentally changed how oncologists think about the tumor microenvironment and immune evasion.
Septic Patient Arginine Deficiency
Sepsis is a syndrome of life-threatening organ dysfunction caused by dysregulated host response to infection. Septic patients show a complex constellation of immune dysfunction including hyper-inflammation early in the course (the cytokine storm), followed by a paradoxical immunosuppression in the protracted phase that increases susceptibility to secondary infection. Arginine depletion contributes to both phases of this dysfunction.
In septic patients, plasma arginine is markedly reduced, typically to 30–50% of normal. The mechanisms are several:
- Massively upregulated arginase activity — circulating arginase derived from neutrophils, MDSCs, and damaged tissue (hemolysis releases erythrocyte arginase) consumes arginine in plasma. Plasma arginase activity can rise 10-fold in severe sepsis.
- Reduced endogenous synthesis — the gut-kidney axis that normally produces arginine from glutamine and citrulline is impaired in sepsis, partly because the intestinal mucosa is damaged (citrulline production is impaired), and partly because acute kidney injury reduces renal capacity for citrulline-to-arginine conversion.
- Increased consumption by activated immune cells — the immune response to infection consumes large amounts of arginine for iNOS-driven NO production by macrophages and neutrophils.
- Reduced dietary intake — critically ill patients often have impaired nutritional intake, compounding the substrate deficit.
The functional consequence of arginine deficiency in sepsis is impaired adaptive immunity (via the CD3-zeta mechanism described above), impaired endothelial function (eNOS substrate depletion contributes to the microvascular dysfunction of septic shock), and impaired wound healing (relevant in trauma sepsis and post-surgical sepsis).
The therapeutic implication is complex. Simple arginine supplementation in unstable septic patients can be harmful (the Bertolini trial, discussed below), likely because adding substrate to upregulated iNOS produces excess NO and worsens hypotension. But arginine substrate restoration is desirable to support adaptive immunity and other functions. Current approaches favor citrulline supplementation in septic patients — citrulline bypasses the upregulated arginase, restores arginine levels gradually, and may avoid the acute hemodynamic concerns of direct arginine loading. Several small trials of citrulline in sepsis have shown promising signals.
ICU Immunonutrition Trials
The history of arginine-containing immunonutrition in the ICU is a cautionary tale about extrapolating from elective surgery to critical illness. The early enthusiasm for arginine-rich enteral formulas was based on data from elective surgical populations, where benefits are clear and consistent. When the same formulas were tested in unstable septic ICU patients, the results were different.
The pivotal negative trial was Bertolini et al. (2003), randomizing 237 ICU patients (including many with sepsis) to standard parenteral nutrition versus parenteral nutrition supplemented with arginine and glutamine. The trial was stopped early when an interim analysis showed significantly higher mortality in the immunonutrition arm. Subsequent meta-analyses by Heyland and colleagues largely confirmed that arginine-rich immunonutrition was associated with increased mortality in the sickest septic patients.
The proposed mechanism was that adding arginine substrate to upregulated iNOS in septic patients produces excess NO, exacerbating septic-shock hypotension and microvascular dysfunction. The clinical consensus that emerged is:
- Elective surgery (stable patients) — arginine-rich immunonutrition is beneficial and should be used in malnourished patients undergoing major procedures
- Trauma without sepsis — arginine-rich immunonutrition appears safe and may be beneficial
- Stable post-op ICU patients — arginine-rich immunonutrition is generally safe
- Severe sepsis and septic shock — arginine-rich formulas should be avoided; standard nutritional support is preferred
- Acute respiratory distress syndrome (ARDS) — the omega-3-rich formulas (without high arginine) have been tested with mixed results; current SCCM and ASPEN guidelines do not strongly recommend specific immunonutrition in ARDS
The current ICU practice in most centers is to use standard high-protein enteral or parenteral nutrition for septic patients, reserving arginine-rich immunonutrition for stable post-operative patients and elective surgical populations. The lessons from the negative sepsis trials have reinforced the importance of distinguishing between different clinical contexts when applying nutritional immunomodulation.
Elective Surgery Immunonutrition
In contrast to the cautionary sepsis literature, the elective surgery literature is consistently positive. Major meta-analyses have shown that perioperative arginine-containing immunonutrition in elective surgical patients reduces infectious complications by 30–50% and reduces length of stay by 1–3 days. The strongest evidence is in upper gastrointestinal cancer surgery and head and neck cancer surgery. This is discussed in detail on the Wound Healing page.
For the immune-function angle specifically, the mechanisms underlying the elective-surgery benefit include:
- Preservation of CD3-zeta and T-cell function — surgery induces a transient post-operative immunosuppression with reduced lymphocyte function. Arginine repletion preserves T-cell activation and reduces the duration of this immunosuppression.
- Enhanced wound antimicrobial defense — via iNOS-driven NO production by wound-recruited macrophages
- Improved gut barrier function — glutamine and arginine together support enterocyte integrity and prevent bacterial translocation that contributes to post-operative infection
- Modulation of the systemic inflammatory response — omega-3 fatty acids in the formula reduce inflammatory eicosanoid production and may dampen the post-surgical SIRS response
The immunological rationale matches the clinical observation: post-operative infection rates are reduced because the immune system is functioning better in the early healing window when most surgical infections originate.
HIV and Chronic Viral Infection
HIV infection produces sustained arginine depletion driven by chronic immune activation, MDSC expansion, and intestinal damage with reduced citrulline production from enterocytes. Plasma arginine is often reduced in untreated HIV patients, and the depletion contributes to the immune dysfunction characteristic of the disease.
Despite the mechanistic rationale, clinical trials of arginine supplementation in HIV have produced mixed results. Some smaller trials have shown improvements in surrogate immunological markers (CD4 count trends, NK function), but larger trials have not demonstrated clinical benefits in terms of opportunistic infection rates or viral load. With the success of modern antiretroviral therapy, the indication for nutritional immunomodulation has receded; ART suppresses viral replication and restores immune function much more reliably than any nutritional intervention.
The arginine-HIV story is mostly of historical interest at this point. The mechanistic principles remain sound, but the clinical context has shifted such that ART is the foundation of treatment and nutritional interventions are supportive at best.
Sickle Cell Disease and Vaso-Occlusive Crisis
Sickle cell disease (SCD) is an inherited hemoglobinopathy in which a single amino acid substitution in the beta-globin chain produces hemoglobin S, which polymerizes under low-oxygen conditions and deforms red blood cells into the characteristic sickle shape. The sickled cells are rigid, fragile, and prone to occluding small vessels, producing the painful vaso-occlusive crises that characterize the disease.
The arginine-SCD connection is striking. Chronic intravascular hemolysis in SCD releases erythrocyte arginase 1 into the plasma, where it depletes circulating arginine. The resulting reduction in NO bioavailability contributes to the pulmonary hypertension, endothelial dysfunction, and possibly the vaso-occlusive crises that drive SCD morbidity. Multiple studies have documented reduced plasma arginine, reduced NO metabolites, and elevated arginase activity in SCD patients.
Clinical trials of arginine supplementation in SCD have shown:
- Reduced opioid requirements during vaso-occlusive crisis — Morris et al. (2013) showed that intravenous L-arginine 200 mg/kg/day given to children hospitalized with vaso-occlusive crisis reduced opioid use by 54% and shortened crisis duration. This was a single-center trial but with a clear positive signal.
- Improvements in pulmonary hypertension — smaller studies have shown that oral arginine supplementation can modestly improve right ventricular function and reduce pulmonary artery pressure in SCD patients with pulmonary hypertension.
- Inconsistent effect on crisis prevention — chronic oral supplementation for prevention of new crises has shown mixed results, with some trials positive and others null.
Current guidelines do not include routine arginine supplementation as standard SCD care, but several SCD specialty centers use arginine adjunctively during acute crises, particularly in pediatric patients. The mechanism is sound and the safety profile is favorable; the question is whether the clinical effect size justifies routine use.
Asthma and the Th2/Arginase Axis
Asthma is a chronic Th2-driven inflammatory disease of the airways characterized by bronchospasm, airway hyperresponsiveness, mucus hypersecretion, and eosinophilic inflammation. The role of arginine in asthma is one of the more surprising chapters in arginine immunology because it does not fit the simple "more arginine is better for immune function" narrative.
In asthma, the Th2 cytokines IL-4 and IL-13 induce arginase 1 expression in airway epithelium and infiltrating macrophages. The upregulated arginase depletes local arginine, reducing substrate available to NOS3 in the bronchial vasculature and to nNOS in the smooth muscle. The result is reduced bronchodilatory NO production, leading to airway hyperresponsiveness. Simultaneously, the arginase-driven ornithine production feeds the polyamine and proline pathways, contributing to airway remodeling and fibrosis.
The therapeutic implication is interesting:
- Arginine supplementation — partially addresses the substrate depletion but also potentially feeds the pathological arginase pathway. Small trials of arginine supplementation in asthma have shown modest improvements in spirometry, but the effect is not large.
- Arginase inhibition — more directly addresses the pathology. Pre-clinical models show that arginase inhibitors reduce airway hyperresponsiveness and improve NO bioavailability. Clinical trials are limited.
- Combination therapy — arginine + arginase inhibition would theoretically restore both substrate supply and NOS access. Not yet clinically tested.
The asthma story illustrates the complexity of arginine biology: in a Th2-dominated inflammatory state, simply adding substrate may not produce the desired NO restoration if the substrate is rapidly diverted by upregulated arginase. The full therapeutic strategy requires addressing both the substrate supply and the enzyme balance.
Key Research Papers
- Rodriguez PC, Ochoa AC (2008). Arginine regulation by myeloid derived suppressor cells and tolerance in cancer: mechanisms and therapeutic perspectives. Immunological Reviews. — PubMed
- Rodriguez PC et al. (2002). Regulation of T cell receptor CD3 zeta chain expression by L-arginine. Journal of Biological Chemistry. — PubMed
- Bronte V, Zanovello P (2005). Regulation of immune responses by L-arginine metabolism. Nature Reviews Immunology. — PubMed
- Geiger R et al. (2016). L-arginine modulates T cell metabolism and enhances survival and anti-tumor activity. Cell. — PubMed
- Bertolini G et al. (2003). Early enteral immunonutrition in patients with severe sepsis: results of an interim analysis of a randomized multicentre clinical trial. Intensive Care Medicine. — PubMed
- Morris CR et al. (2013). A randomized, placebo-controlled trial of arginine therapy for the treatment of children with sickle cell disease hospitalized with vaso-occlusive pain episodes. Haematologica. — PubMed
- Morris SM Jr (2009). Recent advances in arginine metabolism: roles and regulation of the arginases. British Journal of Pharmacology. — PubMed
- Steggerda SM et al. (2017). Inhibition of arginase by CB-1158 blocks myeloid cell-mediated immune suppression in the tumor microenvironment. Journal for ImmunoTherapy of Cancer. — PubMed
- Maarsingh H et al. (2009). Arginase: a key enzyme in the pathophysiology of allergic asthma. Pharmacology and Therapeutics. — PubMed
- Calder PC (2007). Immunonutrition in surgical and critically ill patients. British Journal of Nutrition. — PubMed
- Popovic PJ et al. (2007). Arginine and immunity. Journal of Nutrition. — PubMed
- Munder M (2009). Arginase: an emerging key player in the mammalian immune system. British Journal of Pharmacology. — PubMed
- Luiking YC et al. (2009). Arginine de novo and nitric oxide production in disease states. American Journal of Physiology - Endocrinology and Metabolism. — PubMed
- Heyland DK, Dhaliwal R (2005). Immunonutrition in the critically ill: from old approaches to new paradigms. Intensive Care Medicine. — PubMed
PubMed Topic Searches
- PubMed: Arginine and T-cell CD3-zeta
- PubMed: MDSC arginase in tumors
- PubMed: M1/M2 macrophages and arginine
- PubMed: Arginine deficiency in sepsis
- PubMed: Arginine in sickle cell disease
- PubMed: Immunonutrition in surgery
Connections
- Arginine Overview
- Arginine Benefits Hub
- Arginine for Cardiovascular Health
- Arginine for Erectile Function
- Arginine for Wound Healing
- Glutamine
- Lysine
- All Amino Acids
- Immune Boosting
- Vitamin A for Immune Function
- Vitamin D3
- Zinc
- Diabetes
- ARDS / Critical Illness
- Lysine