Passionflower GABA Modulation
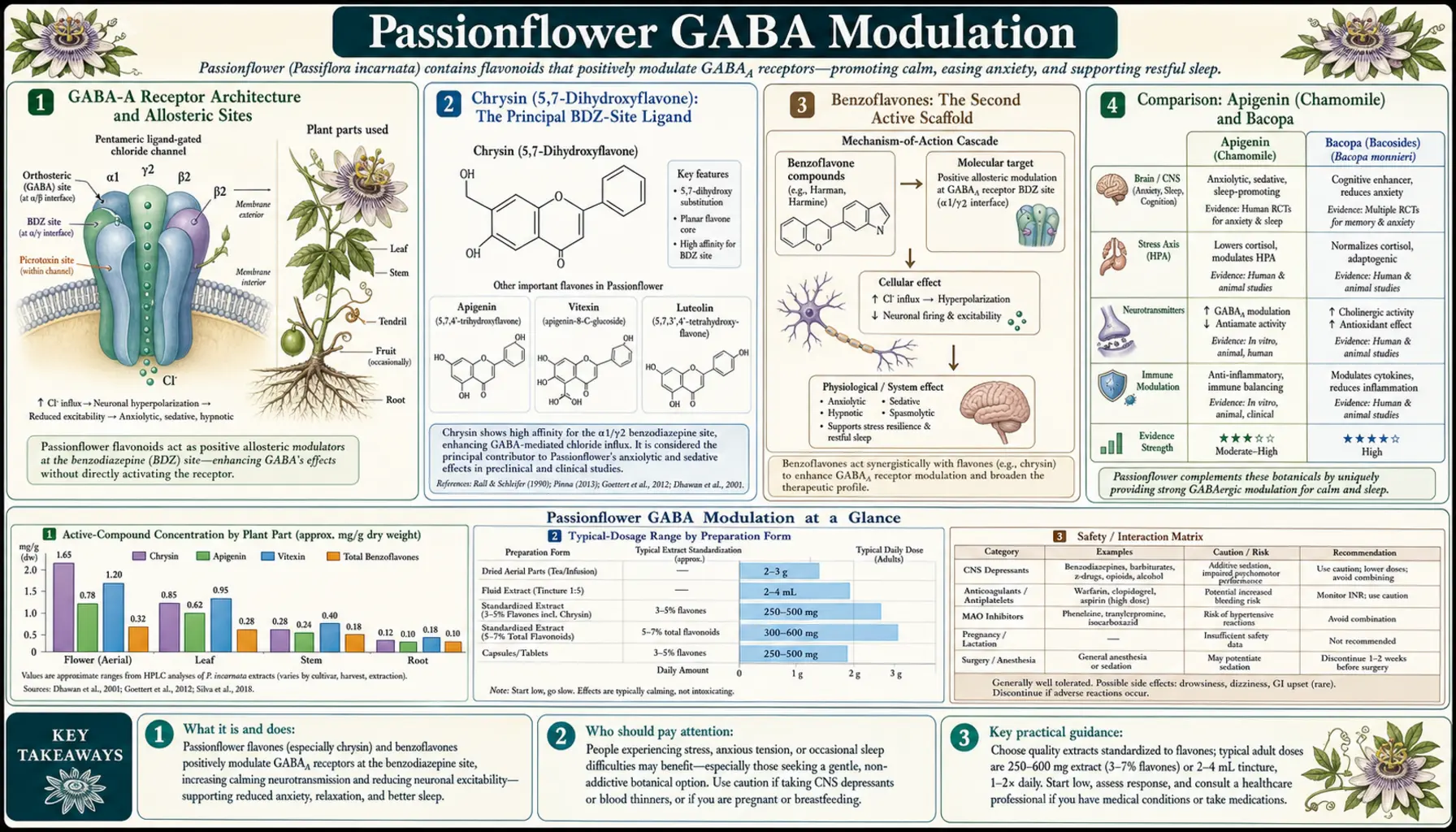
The pharmacology of passionflower centers on the GABA-A receptor, the principal inhibitory ionotropic receptor of the mammalian central nervous system. Wolfman and Medina's 1994 work isolated chrysin (5,7-dihydroxyflavone) from Passiflora coerulea and demonstrated it bound the benzodiazepine site of the GABA-A receptor — the same allosteric pocket where diazepam, lorazepam, alprazolam, and zolpidem exert their effects. Dhawan's 2002 work identified a second active scaffold, a benzoflavone moiety, that also produces anxiolytic effects in rodent models. The combined effect of chrysin, benzoflavones, and several other flavone aglycones is partial-agonist potentiation of GABA-A current, producing modest amplification of endogenous GABAergic inhibition without the powerful clamping effect of full benzodiazepine agonists. Beyond GABA, passionflower also contains trace beta-carboline alkaloids (harman, harmine, harmaline, harmol) that are weak reversible inhibitors of monoamine oxidase A — the basis for theoretical interaction concerns with SSRIs, MAOIs, and tyramine-containing foods. This deep-dive explores the receptor-level pharmacology, comparison with related botanical anxiolytics (apigenin/chamomile, bacopa, valerian), the harman alkaloid story, and what the multi-mechanism profile means for clinical use.
Table of Contents
- GABA-A Receptor Architecture and Allosteric Sites
- Chrysin (5,7-Dihydroxyflavone): The Principal BDZ-Site Ligand
- Benzoflavones: The Second Active Scaffold
- Comparison: Apigenin (Chamomile) and Bacopa
- Non-Benzodiazepine-Site GABA-A Modulation
- Harman Beta-Carboline Alkaloids and MAO-A
- Why Partial Agonism Produces Gentler Effects
- Tolerance, Dependence, and Withdrawal Profile
- The Flumazenil Paradox — Multi-Site Action
- Theoretical and Real Drug Interaction Concerns
- Key Research Papers
- Connections
- Featured Videos
GABA-A Receptor Architecture and Allosteric Sites
The GABA-A receptor is a pentameric chloride-channel ligand-gated ion channel, the dominant fast-inhibitory neurotransmitter receptor in the mammalian central nervous system. Each receptor is built from five protein subunits drawn from a family of 19 known mammalian subunits (alpha1-6, beta1-3, gamma1-3, delta, epsilon, theta, pi, and rho1-3). The most common configuration in adult cortex is 2×alpha, 2×beta, and 1×gamma2, typically with the alpha1 subunit subtype, though other compositions (alpha2, alpha3, alpha5, delta-containing) predominate in specific brain regions and produce distinct pharmacology.
When GABA binds the orthosteric site (located at the alpha-beta subunit interface), the chloride channel opens, chloride flows into the neuron, the neuron hyperpolarizes, and excitability decreases. This is the basic inhibitory function. Multiple allosteric sites on the receptor modulate this response, increasing or decreasing the chloride current produced by a given amount of orthosteric GABA. The pharmacologically important allosteric sites include:
- Benzodiazepine binding site — located at the alpha-gamma subunit interface, requires gamma2 subunit. Positive modulators (diazepam, lorazepam, alprazolam, chrysin, apigenin) potentiate GABA current; negative modulators (beta-carbolines like DMCM) reduce it; antagonists (flumazenil) block other ligands without intrinsic activity
- Barbiturate site — pentobarbital, phenobarbital, propofol. At high doses, can directly open the channel without GABA, which is why barbiturate overdose is lethal in a way benzodiazepine overdose typically is not
- Neurosteroid site — allopregnanolone, THDOC, brexanolone (FDA-approved for postpartum depression). Powerful positive modulators
- Anesthetic site — volatile anesthetics, etomidate, etc.
- Alcohol site(s) — ethanol modulates multiple GABA-A subtypes through poorly defined sites
Passionflower's primary action is at the benzodiazepine site through chrysin and related flavones, with additional activity at non-benzodiazepine-site allosteric pockets through other flavone aglycones (discussed below). This multi-site profile is what distinguishes botanical GABA modulation from single-target pharmaceutical benzodiazepines.
Chrysin (5,7-Dihydroxyflavone): The Principal BDZ-Site Ligand
Chrysin is a flavone (a subclass of flavonoid) with the structural formula 5,7-dihydroxyflavone — a relatively simple flavone with two hydroxyl groups on the A-ring and no substitution on the B-ring. It is widely distributed in plants and is found in honey, propolis, passionflower, and other botanical sources. Medina and colleagues first demonstrated in 1990 that chrysin bound the central benzodiazepine receptor with measurable (though modest) affinity in rat brain membrane assays. Wolfman et al. extended this in 1994 by isolating chrysin from Passiflora coerulea and demonstrating anxiolytic effects in the elevated plus maze in mice, with the effects partially reversed by the benzodiazepine antagonist flumazenil — confirming the BDZ-site mechanism.
The pharmacologic profile of chrysin at the BDZ site is partial-agonist with low-to-moderate affinity. Compared to a full agonist like diazepam (Ki ~10 nM), chrysin's affinity is in the micromolar range, and even at saturating concentrations chrysin produces only a fraction of the maximal positive modulation that diazepam produces. This is exactly the pharmacology one would design for a gentle anxiolytic: enough BDZ-site occupation to amplify GABAergic inhibition modestly, but never enough to clamp the chloride channel and produce the cognitive impairment, motor incoordination, amnestic effect, and dependence liability of full benzodiazepine agonists.
Bioavailability is a significant constraint. Chrysin has poor oral bioavailability in humans (typically < 1% reported in pharmacokinetic studies), with extensive first-pass metabolism producing chrysin glucuronide and sulfate conjugates. This raises the legitimate question of whether enough free chrysin reaches the brain after an oral passionflower dose to account for the clinical effect. The answer is probably partly chrysin and partly the other actives discussed below — benzoflavones, additional flavone aglycones, and the still-undefined contribution of the harman alkaloids. The whole-extract is more active than isolated chrysin would predict, consistent with multi-component synergy.
Benzoflavones: The Second Active Scaffold
Dhawan and Sharma at Punjab University identified a tricyclic benzoflavone moiety in Passiflora incarnata methanol extract through bioassay-guided fractionation in a series of papers from 1999-2003. The benzoflavone is structurally distinct from chrysin — an additional fused benzene ring extends the flavone scaffold — and produced anxiolytic effects in the elevated plus maze at doses lower than the parent extract on a milligram basis, suggesting it accounts for a substantial fraction of the overall Passiflora incarnata activity.
The benzoflavone moiety has been the subject of additional investigation:
- Anxiolytic effect in rodent models confirmed by multiple independent groups
- Reversal of morphine tolerance and dependence in mice (Dhawan 2002)
- Reversal of nicotine tolerance and dependence in mice (Dhawan 2002)
- Reversal of cannabinoid (delta-9-THC) tolerance in mice (Dhawan 2002)
- Benefit on aged-rat reproductive parameters (Dhawan 2003)
The mechanism by which the benzoflavone produces these diverse effects is not fully characterized but likely involves modulation of multiple receptor systems including the GABA-A receptor, possibly the cannabinoid CB1 receptor (theorized but not confirmed), and downstream effects on stress-hormone axes. The reversal-of-tolerance effects across opiate, nicotine, and cannabinoid models is a particularly intriguing common-pathway finding that has guided the use of passionflower in addiction medicine.
Standardization of passionflower extracts to benzoflavone content (rather than chrysin or vitexin) has been proposed by some authors as more clinically meaningful, but commercial standardization remains heterogeneous — some products specify vitexin equivalents, others specify total flavonoid content, others use no standardization at all.
Comparison: Apigenin (Chamomile) and Bacopa
Several other widely used botanical anxiolytics work through the same GABA-A benzodiazepine binding site as chrysin, with structurally similar flavone scaffolds:
- Apigenin (chamomile) — structurally chrysin + a 4'-hydroxyl on the B-ring (so apigenin = 4',5,7-trihydroxyflavone). Apigenin binds the benzodiazepine site with similar affinity to chrysin and produces similar partial-agonist effects. Chamomile's anxiolytic effect is documented in the Amsterdam 2009 GAD trial. Apigenin is generally considered slightly milder than chrysin in animal models, consistent with chamomile's clinical reputation as gentler than passionflower
- Bacopasides (Bacopa monnieri) — the triterpene saponins of bacopa modulate GABA-A receptors among multiple other targets (including serotonin, dopamine, and cholinergic systems). Bacopa is more often used for cognitive enhancement than anxiolysis specifically, but the GABA mechanism contributes to its calming side effect
- Honokiol and magnolol (Magnolia officinalis) — biphenolic compounds that modulate GABA-A through a site distinct from the benzodiazepine site. Stronger sedative effect than passionflower
- Skullcap (Scutellaria baicalensis) — the flavone baicalein has BDZ-site affinity. Used in traditional Chinese medicine for anxiety
- Hops (Humulus lupulus) — 2-methyl-3-buten-2-ol and related compounds at GABA-A site. Often combined with passionflower and valerian in sleep formulations
- Wild lettuce (Lactuca virosa) — lactucin and lactucopicrin produce mild sedation through poorly characterized GABA modulation
The therapeutic implication of this large family of GABA-A modulating flavones is that combination herbal formulations can layer multiple low-affinity ligands to produce additive effect without redundancy. A formulation containing passionflower + chamomile + skullcap may produce more robust anxiolysis than any single herb alone at comparable total dose, while maintaining the cognitive-preservation profile of partial-agonist pharmacology.
Non-Benzodiazepine-Site GABA-A Modulation
An important pharmacologic nuance: not all of passionflower's GABA-A modulating activity is at the benzodiazepine binding site. The Appel 2011 work using cloned recombinant GABA-A receptors expressed in oocytes demonstrated that Passiflora incarnata extract potentiated GABA currents through both BDZ-site-dependent and BDZ-site-independent mechanisms. The independent component was not blocked by flumazenil, indicating action at a different allosteric site on the receptor.
The Elsas 2010 study using hippocampal neurons in culture extended this finding by showing that passionflower extract produced GABA-A currents that were partially flumazenil-resistant, consistent with a multi-site mechanism. The identity of the non-BDZ allosteric site is not fully characterized but may involve an interaction with the alpha subunit or a novel binding pocket.
The clinical relevance: a patient who is taking flumazenil (used in the emergency department to reverse benzodiazepine overdose, or rarely chronically to manage hepatic encephalopathy) would not have the full anxiolytic effect of a pure BDZ-site ligand blocked, only the BDZ-site component. The flumazenil-resistant component of passionflower's activity should remain intact. This is a niche but real distinction that contributes to passionflower's distinctive pharmacologic profile.
Harman Beta-Carboline Alkaloids and MAO-A
Beyond flavonoids, Passiflora incarnata contains trace amounts of beta-carboline alkaloids in the harman family — harman (1-methyl-beta-carboline), harmine (1-methyl-7-methoxy-beta-carboline), harmaline (the dihydro analog of harmine), and harmol (the 7-hydroxy analog). These compounds are weak reversible inhibitors of monoamine oxidase A (MAO-A), the enzyme that catabolizes serotonin, norepinephrine, and dietary tyramine.
The harman alkaloid content in passionflower is low — typically reported in the range of 0.01-0.1% of dried herb weight, with substantial variation across cultivars, growing conditions, and harvest timing. The pharmacologically relevant concentrations achieved with normal therapeutic passionflower doses are well below those required to produce clinically meaningful MAO inhibition in vivo. For comparison, the same beta-carboline alkaloids in much higher concentration in the Amazonian vine Banisteriopsis caapi (ayahuasca) do produce potent MAO-A inhibition, but at orders-of-magnitude higher concentration than is achieved with normal passionflower dosing.
That said, the presence of these alkaloids creates a theoretical concern for drug interactions:
- SSRIs and SNRIs — theoretical risk of serotonin syndrome from additive serotonin elevation. Clinical reports with passionflower alone are essentially absent, but the theoretical interaction is worth mentioning when patients are on high-dose SSRIs (escitalopram > 20 mg, sertraline > 150 mg, fluoxetine > 60 mg)
- Prescribed MAOIs (phenelzine, tranylcypromine, selegiline) — theoretical additive MAO inhibition. Avoid in patients on prescribed MAOIs as a precautionary practice, even though the actual MAO-inhibitor contribution of passionflower is minimal
- Tyramine-containing foods — the classic MAOI dietary interaction. Not a real concern at normal passionflower doses but worth understanding the underlying mechanism
- Triptans (sumatriptan, etc.) — theoretical serotonin syndrome risk; clinically not documented
- Tramadol, meperidine — theoretical risk; clinically not documented
The Avula 2012 work quantified beta-carboline and flavonoid content across multiple Passiflora species and commercial products by HPLC, providing the analytical basis for any quality-control standardization. Most clinical concerns about MAO-mediated interactions are theoretical rather than documented in practice; the actual clinical safety record of passionflower in routine use is excellent.
Why Partial Agonism Produces Gentler Effects
The partial-agonist character of chrysin and benzoflavones at the GABA-A benzodiazepine site is the central mechanistic explanation for passionflower's gentler clinical profile relative to prescription benzodiazepines. The pharmacologic principle:
- Full agonist (diazepam, alprazolam, lorazepam) — produces maximal positive modulation of GABA-A current at saturating concentration. The receptor functions at its maximum gain. At therapeutic doses, occupancy of the BDZ site is high (50-90% in PET imaging studies), producing strong global GABAergic inhibition. The result is robust anxiolysis but also cognitive impairment, motor incoordination, amnesia, sedation, and (with chronic use) downregulation of GABA-A receptors producing tolerance and physiologic dependence
- Partial agonist (chrysin, benzoflavones) — produces only a fraction of the maximal positive modulation, even at saturating concentration. Plus, low affinity means that at clinically achievable concentrations, BDZ-site occupancy is modest (likely 10-30% in animal models). The combined effect is modest amplification of basal GABA-A current — enough to reduce pathologic anxiety but insufficient to clamp arousal, cognition, or motor coordination
The partial-agonist profile also has a self-limiting ceiling on effect: increasing the dose beyond a certain point does not produce proportionally more anxiolysis because the inherent intrinsic activity of the molecule is limited. This is why passionflower has an excellent overdose safety profile — it is essentially impossible to produce life-threatening respiratory depression with passionflower alone, in contrast to benzodiazepines (especially in combination with opioids or alcohol).
The trade-off is that severe acute anxiety, panic attacks, status epilepticus, or severe insomnia — situations that require maximal GABA-A potentiation — will not be adequately treated by passionflower or any partial-agonist botanical. Full benzodiazepine agonism remains the standard for these acute indications.
Tolerance, Dependence, and Withdrawal Profile
The published clinical and observational evidence base for passionflower does not document significant tolerance development, physiologic dependence, or withdrawal syndrome with chronic use. This is mechanistically consistent with the low-occupancy, partial-agonist binding profile:
- Tolerance — full benzodiazepine agonists produce tolerance through GABA-A receptor downregulation and subunit composition shifts in chronic use. The signaling required to drive these adaptive changes is proportional to receptor occupancy and activation. Partial agonists at low occupancy do not generate enough adaptive signaling to produce meaningful tolerance over weeks-to-months of chronic dosing
- Physiologic dependence — the molecular basis of benzodiazepine dependence is the same downregulation that drives tolerance. Without downregulation, abrupt discontinuation does not produce a rebound hyperexcitable state. Passionflower discontinuation does not produce documented withdrawal in clinical use
- Psychological dependence — possible in patients who have come to associate the ritual of taking passionflower with sleep onset or anxiety relief. Generally mild and reversible with conscious tapering
This favorable profile is one of the main reasons passionflower has a legitimate role in benzodiazepine tapering protocols, where its mild GABA-A potentiation can help ease the rebound anxiety and insomnia of taper without becoming a substitute dependence target.
The Flumazenil Paradox — Multi-Site Action
Flumazenil is a pharmaceutical antagonist at the GABA-A benzodiazepine binding site, used in the emergency department to reverse benzodiazepine overdose. A purely BDZ-site ligand should have its effect completely blocked by flumazenil pretreatment.
In animal models, flumazenil partially but not completely blocks the anxiolytic effect of passionflower extract. The Wolfman 1994 work with isolated chrysin found flumazenil reversed most of chrysin's effect, consistent with pure BDZ-site action. But the Appel 2011 work with whole extract using recombinant GABA-A receptors found a substantial flumazenil-resistant component, and the Elsas 2010 work in hippocampal neurons confirmed this.
The interpretation: passionflower has multiple active components, and not all of them work through the benzodiazepine binding site. The flavonoid pool likely includes ligands at multiple distinct allosteric sites on the GABA-A receptor. The benzoflavones may work at a different site than chrysin. There may also be modest direct GABA-mimetic activity from one or more components. The whole-extract pharmacology is therefore more complex than isolated chrysin would predict.
Clinically, this means passionflower's effect cannot be fully reversed by flumazenil — a niche but real distinction that contributes to its distinctive profile. It also reinforces the general principle that whole-plant botanical pharmacology is rarely reducible to a single isolated active compound.
Theoretical and Real Drug Interaction Concerns
- Benzodiazepines and Z-drugs — theoretical additive GABA-A potentiation. Practically, the additive effect is mild because passionflower's contribution is modest. Useful in benzodiazepine tapering but use caution combining with full agonists in elderly or frail patients
- Alcohol — additive CNS depression via GABA-A modulation and other mechanisms. Avoid significant alcohol intake when taking passionflower, especially for driving or operating machinery
- Opioids — theoretical additive sedation. Useful in opioid withdrawal protocols (see Withdrawal & Addiction Support) under supervision, but avoid combining recreational opioid use with passionflower at home
- Sedating antihistamines (diphenhydramine, doxylamine) — additive sedation
- Sedating antidepressants (trazodone, mirtazapine, doxepin) — additive sedation; can sometimes be used productively for stubborn insomnia but monitor for excessive morning sedation
- SSRIs/SNRIs — theoretical serotonin elevation from harman MAO-A inhibition. Clinically not a documented concern at normal doses
- Prescribed MAOIs (rare) — avoid combination as precaution
- Warfarin and other anticoagulants — some sources suggest theoretical platelet effects from passionflower flavonoids; clinical reports of bleeding are absent. Mention to physician if anticoagulated
- General anesthesia — standard practice is to stop herbal sedatives 1-2 weeks before scheduled surgery as precaution. Probably overcautious but the standard recommendation
- Pregnancy — insufficient safety data plus theoretical uterine stimulant effects from harman alkaloids. Avoid in pregnancy
- Breastfeeding — limited data; avoid or use minimum effective dose with infant monitoring
Key Research Papers
- Medina JH, Paladini AC, Wolfman C, Levi de Stein M, Calvo D, Diaz LE, Pena C (1990). Chrysin (5,7-di-OH-flavone), a naturally occurring ligand for benzodiazepine receptors, with anticonvulsant properties. Biochemical Pharmacology, 40(10), 2227-2231. — PubMed
- Wolfman C, Viola H, Paladini A, Dajas F, Medina JH (1994). Possible anxiolytic effects of chrysin, a central benzodiazepine receptor ligand isolated from Passiflora coerulea. Pharmacology Biochemistry and Behavior, 47(1), 1-4. — PubMed
- Dhawan K, Kumar S, Sharma A (2001). Anti-anxiety studies on extracts of Passiflora incarnata Linneaus. Journal of Ethnopharmacology, 78(2-3), 165-170. — PubMed
- Dhawan K, Dhawan S, Sharma A (2004). Passiflora: a review update. Journal of Ethnopharmacology, 94(1), 1-23. — PubMed
- Marder M, Paladini AC (2002). GABA-A-receptor ligands of flavonoid structure. Current Topics in Medicinal Chemistry, 2(8), 853-867. — PubMed
- Appel K, Rose T, Fiebich B, Kammler T, Hoffmann C, Weiss G (2011). Modulation of the gamma-aminobutyric acid (GABA) system by Passiflora incarnata L. Phytotherapy Research, 25(6), 838-843. — PubMed
- Elsas SM, Rossi DJ, Raber J, White G, Seeley CA, Gregory WL, Mohr C, Pfankuch T, Soumyanath A (2010). Passiflora incarnata L. (passionflower) extracts elicit GABA currents in hippocampal neurons in vitro, and show anxiogenic and anticonvulsant effects in vivo, varying with extraction method. Phytomedicine, 17(12), 940-949. — PubMed
- Salgueiro JB, Ardenghi P, Dias M, Ferreira MB, Izquierdo I, Medina JH (1997). Anxiolytic natural and synthetic flavonoid ligands of the central benzodiazepine receptor have no effect on memory tasks in rats. Pharmacology Biochemistry and Behavior, 58(4), 887-891. — PubMed
- Avula B, Wang YH, Rumalla CS, Smillie TJ, Khan IA (2012). Quantitative determination of indole and beta-carboline alkaloids and flavonoids of Passiflora species using UHPLC. Acta Chromatographica, 24(1), 75-90. — PubMed
- Sigel E, Steinmann ME (2012). Structure, function, and modulation of GABA-A receptors. Journal of Biological Chemistry, 287(48), 40224-40231. — PubMed
- Hanrahan JR, Chebib M, Johnston GA (2011). Flavonoid modulation of GABA(A) receptors. British Journal of Pharmacology, 163(2), 234-245. — PubMed
- Johnston GA (2015). Flavonoid nutraceuticals and ionotropic receptors for the inhibitory neurotransmitter GABA. Neurochemistry International, 89, 120-125. — PubMed
PubMed Topic Searches
- PubMed: Chrysin BDZ receptor
- PubMed: Passiflora benzoflavone
- PubMed: Flavonoid GABA-A partial agonist
- PubMed: Harman MAO-A inhibition
- PubMed: Passiflora flumazenil reversal
Connections
- Passionflower Overview
- Passionflower Benefits Hub
- Passionflower for Anxiety Relief
- Passionflower for Sleep Quality
- Passionflower for Withdrawal
- Chamomile (Apigenin)
- Valerian (Valerenic Acid)
- Lavender (Linalool)
- Lemon Balm (Rosmarinic Acid)
- Kava (Kavalactones)
- Magnesium (GABA Cofactor)
- Anxiety
- Insomnia
- Natural Anxiety Relief
- Stress Management
- All Herbs