Isoleucine for Hemoglobin and Wound Healing
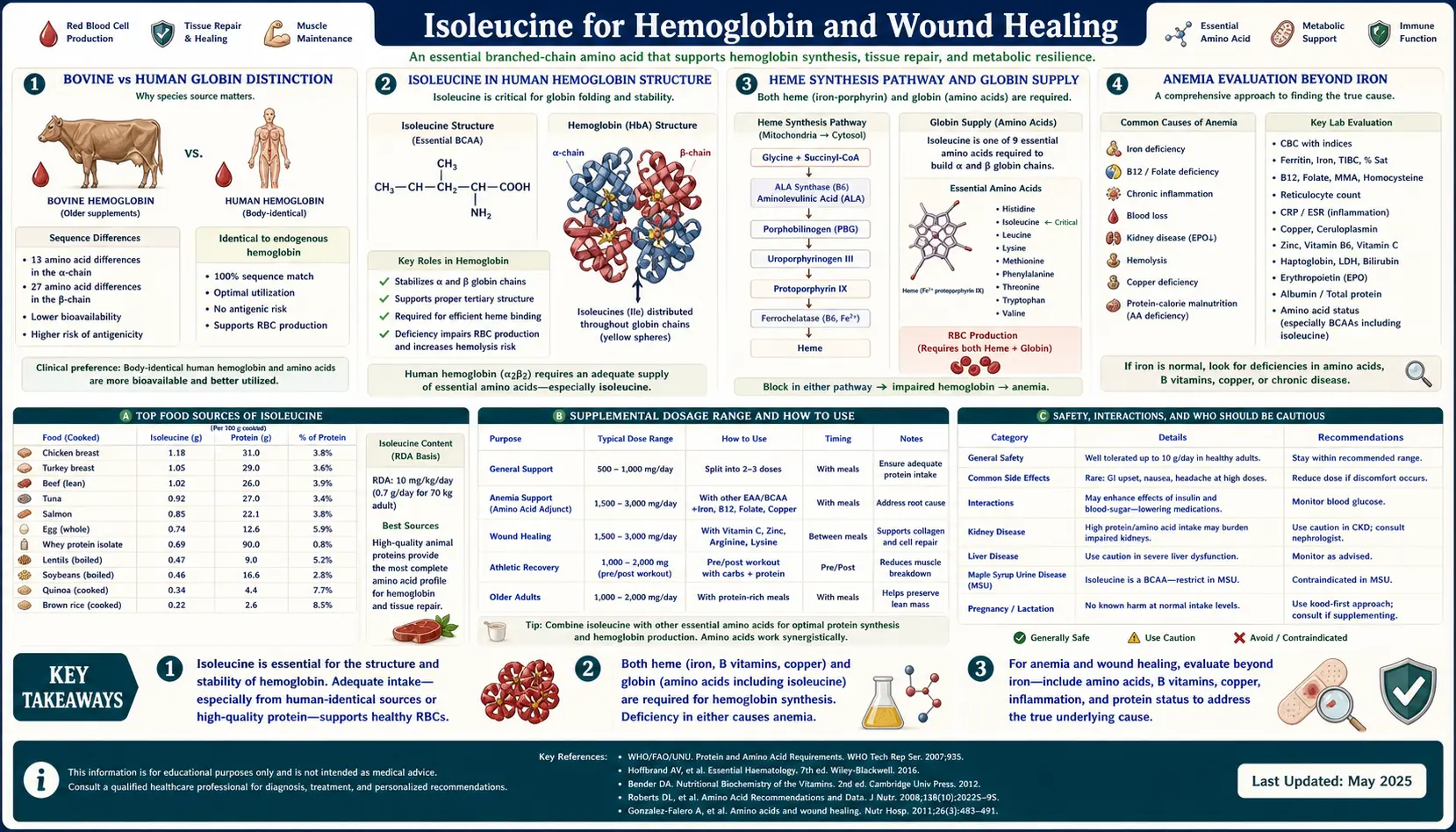
Of the four deep-dive pages in this Benefits cluster, the hemoglobin and wound-healing topic is the most underappreciated — partly because of a single biological fact that gets buried in nearly every introductory biochemistry textbook: bovine globin lacks isoleucine. Specifically, the alpha and beta chains of bovine hemoglobin contain no isoleucine residues, while human hemoglobin contains isoleucine at multiple critical positions (alpha chain residues 6 and 17; beta chain residues 75 and 137). This is a classic textbook example of how minor differences in amino acid composition between species reveal which residues are functionally essential versus dispensable for a protein's core operation. For humans, isoleucine is required for proper hemoglobin synthesis, and isoleucine status directly affects red blood cell production. Beyond hemoglobin, isoleucine plays a documented role in beta-defensin upregulation (a key innate immune defense at mucosal surfaces) and in the protein-synthesis-intensive phases of wound healing. This deep-dive walks through the bovine-vs-human distinction, the molecular role of isoleucine in heme protein structure, the beta-defensin mechanism, and the clinical relevance for anemia evaluation, post-surgical recovery, and chronic wound care.
Table of Contents
- The Bovine vs Human Globin Distinction
- Isoleucine in Human Hemoglobin Structure
- The Heme Synthesis Pathway and Globin Supply
- Anemia Evaluation Beyond Iron
- Myoglobin and the Cytochrome Family
- Beta-Defensins and Innate Mucosal Immunity
- Wound Healing — the Three-Phase Protein Demand
- Surgical and Burn Recovery Applications
- Chronic Wounds — Diabetic Ulcers and Pressure Sores
- Dietary Strategy for Hemoglobin and Healing Support
- Cautions
- Key Research Papers
- Connections
- Featured Videos
The Bovine vs Human Globin Distinction
Open almost any introductory protein biochemistry textbook and somewhere in the chapter on hemoglobin you will find a sequence comparison between human and bovine globin chains. The comparison is included as a classic teaching example for two reasons: it illustrates how proteins can perform the same function (oxygen binding and release) despite differences in primary sequence, and it raises the question of which amino acid residues are functionally essential versus structurally dispensable.
One specific finding from the comparison is consistently noted: bovine alpha-globin and beta-globin chains contain zero isoleucine residues. The cow makes its entire hemoglobin protein using the other nineteen amino acids, with isoleucine absent from the chain. Human alpha-globin, by contrast, contains isoleucine at two positions (residues 6 and 17 in the mature 141-residue chain); human beta-globin contains isoleucine at residues 75 and 137 in the mature 146-residue chain. The isoleucine residues participate in the hydrophobic core of the folded globin tertiary structure, contributing to the stability of the alpha-helical bundle that creates the heme-binding pocket.
The bovine-vs-human contrast is presented in textbooks as evidence that isoleucine, while present in human hemoglobin, is not strictly required for the oxygen-binding chemistry itself — bovine hemoglobin functions perfectly well without it, by substituting other hydrophobic residues (typically leucine or valine) at the equivalent positions. What it does demonstrate is that the human hemoglobin synthesis machinery, having evolved with isoleucine in the sequence, depends on isoleucine availability to produce normally folded, functional human hemoglobin. If isoleucine is insufficient during periods of high erythropoietic demand (recovery from blood loss, response to high altitude, post-partum recovery), human red blood cell production is impaired even when iron, B12, folate, and other "classical" hematinic nutrients are adequate.
This is the practical reason isoleucine appears in lists of nutrients required for hemoglobin synthesis: not because it is universally essential for oxygen-binding chemistry across species, but because the human-specific globin sequence requires isoleucine, and isoleucine deficiency can therefore present clinically as a nutritional anemia that does not respond to iron supplementation alone.
Isoleucine in Human Hemoglobin Structure
Human adult hemoglobin (HbA) is a tetramer of two alpha-globin chains and two beta-globin chains, each chain folded into a compact globin fold consisting of eight alpha-helices labeled A through H. The four chains together create the cooperative oxygen-binding behavior (the sigmoidal oxygen-hemoglobin dissociation curve) that allows efficient oxygen loading in the lungs and oxygen release in peripheral tissues.
The isoleucine residues in human globin are not surface-exposed; they are buried in the hydrophobic interior of the folded protein, where they participate in the structural stability of the globin fold. Specifically:
- Alpha-globin Ile-6 sits in the A-helix and participates in the packing of the A and H helices against one another. The hydrophobic isoleucine side chain fills a cavity between the helices and stabilizes the orientation that creates the heme-binding pocket on the opposite side of the molecule.
- Alpha-globin Ile-17 participates in the A-B helix interface, with similar structural-packing function.
- Beta-globin Ile-75 is in the E-F corner region (between the E and F helices), close to the proximal histidine (His-92) that coordinates the heme iron. Substitutions at this position produce known hemoglobinopathies, indicating the residue's functional importance.
- Beta-globin Ile-137 is in the H-helix and participates in subunit-subunit contact with alpha-globin in the formation of the tetramer.
The hydrophobic packing role of isoleucine is functionally substitutable by leucine or valine (both also branched-chain hydrophobic amino acids), which is why bovine hemoglobin can function without isoleucine — the bovine sequence uses leucine or valine at the equivalent positions. But once the human gene encodes isoleucine codons at those positions, the cellular protein synthesis machinery must have isoleucine available to translate those codons. Substitution at the synthesis step is not possible; the ribosome will not insert leucine where the codon specifies isoleucine. If isoleucine is unavailable, translation stalls, the integrated stress response is triggered, and erythroid cells reduce globin production.
The Heme Synthesis Pathway and Globin Supply
Hemoglobin synthesis in developing red blood cells (erythroblasts) requires the coordinated production of heme (the iron-containing porphyrin cofactor) and globin (the protein chains that bind the heme). Heme synthesis takes place partly in the mitochondria and partly in the cytoplasm of the erythroblast, in an eight-step enzymatic pathway that requires iron, succinyl-CoA (which provides the carbon backbone of the porphyrin ring), glycine, and several B vitamins as cofactors.
Globin synthesis happens on cytoplasmic ribosomes and requires the full complement of the twenty amino acids that comprise the globin chain sequence. The two processes are tightly coupled — mature hemoglobin requires both heme and globin in stoichiometric balance, and isolated production of either component triggers feedback that limits the other.
For isoleucine specifically, its role in hemoglobin production is at the globin-synthesis step. Erythroblasts producing huge quantities of alpha and beta globin chains during the late stages of erythroid maturation have enormous translation demands — a single mature human red blood cell contains approximately 270 million hemoglobin molecules, each requiring four globin chains, meaning roughly one billion globin chains per cell. The cellular isoleucine pool is consumed rapidly during this synthesis burst, and the surrounding tissue must supply isoleucine continuously to keep up.
In severe protein-energy malnutrition (kwashiorkor and marasmus), one of the hematological findings is normocytic normochromic anemia — not the microcytic hypochromic anemia of iron deficiency, but a different pattern reflecting impaired globin production despite adequate iron stores. The mechanism is generalized amino acid insufficiency limiting globin synthesis. Isoleucine is one of the essential amino acids in this insufficiency, alongside lysine, threonine, leucine, valine, methionine, phenylalanine, tryptophan, histidine, and (in infants) tyrosine.
Anemia Evaluation Beyond Iron
The standard clinical workup for anemia focuses on the classic hematinic deficiencies: iron (microcytic hypochromic pattern), B12 (macrocytic, often with neurological signs), folate (macrocytic, megaloblastic bone marrow), and the inflammatory anemia of chronic disease. Protein and amino acid deficiency is less commonly considered in the differential, partly because it is relatively rare in developed-country populations and partly because the laboratory tests for amino acid status are not part of routine anemia workup.
When anemia evaluation should include consideration of protein and amino acid (including isoleucine) status:
- Anemia that does not respond to iron repletion. A patient with iron deficiency anemia who is given adequate oral or IV iron and whose ferritin and transferrin saturation normalize but whose hemoglobin does not rise — the anemia is iron-replete but persisting. Causes include chronic inflammation, occult bleeding, hemolysis, and (less commonly) protein-amino-acid insufficiency.
- Strict vegan with inadequate dietary planning. A vegan eating mostly fruit and refined grains without deliberate inclusion of legumes, soy, nuts, seeds, and complete plant-protein combinations can develop subtle essential amino acid deficiencies, including isoleucine. The anemia is typically mild and normocytic; serum amino acid profiling can document the deficiency.
- Malabsorption. Crohn's disease, celiac disease, gastric bypass, chronic pancreatitis, and short-bowel syndrome can produce generalized protein and micronutrient malabsorption. Amino acid panels can be diagnostic.
- Post-bariatric surgery patients. Both protein malabsorption and reduced food intake contribute to amino acid deficiency. Routine annual amino acid screening is reasonable in post-RYGB and post-DS patients.
- Severe protein-energy malnutrition. In international medicine and refugee/famine contexts, kwashiorkor and marasmus present with anemia that is amino-acid-limited rather than iron-limited. Repletion requires complete protein, not iron supplementation alone.
The serum amino acid panel (typically a 20-30 amino acid quantitative analysis) is the most direct tool for diagnosing isoleucine and other essential amino acid deficiency. Plasma isoleucine concentration below approximately 30 micromolar (normal: 50-100 micromolar) indicates a functional deficiency that can affect hemoglobin synthesis and other protein-synthetic processes.
For the broader clinical picture of anemia evaluation, see our Anemia page.
Myoglobin and the Cytochrome Family
Hemoglobin is the most quantitatively important heme protein in the body, but it is not the only one. Other heme-containing proteins that require isoleucine for proper synthesis include:
- Myoglobin — the muscle-localized oxygen storage protein. Myoglobin is a single-chain globin (153 amino acids) structurally homologous to hemoglobin subunits. Human myoglobin contains isoleucine residues that contribute to the same hydrophobic-packing function as in hemoglobin. Myoglobin is concentrated in slow-twitch muscle fibers and gives red meat its color (and its iron content). Insufficient myoglobin reduces the oxygen-carrying capacity of muscle tissue itself, contributing to exercise intolerance and fatigue.
- Cytochrome c and the other cytochromes of the mitochondrial electron transport chain — these heme proteins are essential for cellular oxidative phosphorylation (ATP production). Cytochrome c is a small protein (104 amino acids) containing isoleucine residues that participate in the hydrophobic core. Reduced cytochrome production impairs cellular energy production across all tissues.
- Cytochrome P450 enzymes — the large family of heme-containing enzymes responsible for phase I drug and xenobiotic metabolism in the liver. The various P450 isoforms contain isoleucine residues at multiple positions in their catalytic domains. Reduced P450 capacity impairs the liver's ability to metabolize drugs, hormones, and toxins.
- Catalase — the antioxidant enzyme that converts hydrogen peroxide to water and oxygen. Catalase is a tetramer of 506-residue subunits, each containing one heme. Reduced catalase activity allows hydrogen peroxide accumulation, contributing to oxidative stress.
- Peroxidases — including myeloperoxidase (in neutrophils, contributing to bacterial killing) and lactoperoxidase (in saliva and milk, with antimicrobial function).
The collective effect of isoleucine insufficiency on this family of heme proteins is multisystem: reduced oxygen-carrying capacity in blood and muscle, impaired mitochondrial energy production, reduced detoxification capacity, increased oxidative stress, and impaired innate immune function. This is one reason that severe protein-energy malnutrition produces such a global clinical picture of fatigue, infection susceptibility, and metabolic dysfunction — it is not just hemoglobin that suffers, but the entire heme-protein ecosystem.
Beta-Defensins and Innate Mucosal Immunity
Beta-defensins are small (28-42 amino acid) cysteine-rich antimicrobial peptides secreted by epithelial cells at mucosal surfaces (gut, lung, skin, urogenital tract). They are part of the innate immune system's first-line defense, killing bacteria, fungi, enveloped viruses, and some protozoa by disrupting microbial membranes. Unlike antibodies, which take days to weeks to develop after pathogen exposure, beta-defensins are constitutively produced or rapidly upregulated within hours of infection or epithelial damage.
The interesting connection to isoleucine: research from the early 2000s (notably the work of Charles Bevins, Andre Ouellette, and others) showed that BCAA availability — particularly isoleucine — regulates beta-defensin gene expression in epithelial tissues. Isoleucine appears to act as a signaling molecule that licenses transcription of the human beta-defensin 2 (hBD-2) and hBD-3 genes in epithelial cells. The exact molecular mechanism is still being characterized, but the functional result is clear: increased isoleucine availability increases epithelial beta-defensin production, increasing the innate antimicrobial barrier at mucosal surfaces.
This has potential therapeutic relevance. Patients with chronic mucosal infections (recurrent UTIs, chronic sinusitis, chronic gastrointestinal infection) may benefit from optimization of BCAA intake to support epithelial beta-defensin production. Animal studies have shown that dietary isoleucine supplementation enhances mucosal antimicrobial defense and reduces colonization by various enteric pathogens. The human clinical evidence is more preliminary — isoleucine has not yet emerged as a standard nutritional intervention for any specific infectious indication — but the mechanism is interesting and may support recommendations for adequate protein intake in patients with chronic mucosal infection burden.
The beta-defensin mechanism is also a candidate explanation for why protein-malnourished populations show such dramatic increases in mucosal infection (diarrhea, pneumonia, otitis media). In addition to the well-described effects of malnutrition on adaptive immunity, the innate antimicrobial peptide defense at mucosal surfaces is also compromised when amino acid substrate is inadequate.
Wound Healing — the Three-Phase Protein Demand
Wound healing proceeds through three overlapping phases, each with distinct protein synthesis demands that can be limited by isoleucine availability:
Inflammatory phase (days 0-3). Immediately after injury, the wound bed is populated by neutrophils and macrophages that clear debris and dead tissue, and by activated platelets that initiate coagulation. The inflammatory phase requires synthesis of cytokines (TNF-alpha, IL-1, IL-6), chemokines (IL-8, MCP-1), complement components, antimicrobial peptides (defensins, including the beta-defensins discussed above), and the cellular machinery for phagocytosis and respiratory burst. All of this requires amino acid substrate, including isoleucine.
Proliferative phase (days 3-21). Fibroblasts proliferate at the wound site and synthesize large quantities of collagen (the dominant structural protein of connective tissue) and other extracellular matrix proteins (fibronectin, elastin, proteoglycans). Endothelial cells proliferate and form new capillaries (angiogenesis). Keratinocytes re-epithelialize the wound surface. This phase has the highest sustained protein synthesis demand of the healing process. Collagen is roughly 6% isoleucine by mass; if isoleucine is limiting, collagen production slows and wound matrix formation is delayed.
Remodeling phase (week 3 through months to years). The initial type III collagen of the proliferative phase is gradually replaced by stronger type I collagen, with crosslinks formed by lysyl oxidase (a copper-dependent enzyme — see our Copper page). The wound contracts and the scar matures. This phase has lower acute protein synthesis demand but requires sustained substrate availability over months. Inadequate protein intake during this phase results in weak, fragile scars that are prone to re-injury.
Across all three phases, isoleucine's role is as a building block for the proteins being synthesized. Its anti-catabolic effect in muscle also matters — during the inflammatory and proliferative phases, the body's metabolic state can shift toward net catabolism (driven by elevated cortisol, inflammatory cytokines, and the energy demand of the healing process), and muscle protein breakdown can be a source of amino acids for the wound. Adequate dietary BCAA intake helps spare muscle protein and shift the source of healing amino acids toward dietary intake.
Surgical and Burn Recovery Applications
Two clinical contexts where amino acid (including isoleucine) supplementation has been most systematically studied:
Post-surgical recovery. Major surgery (orthopedic joint replacement, cardiac surgery, abdominal surgery, organ transplantation) produces a hypermetabolic catabolic state that can persist for days to weeks. Net protein breakdown accelerates, muscle mass is lost, and wound healing competes with muscle preservation for available amino acids. Standard enhanced recovery after surgery (ERAS) protocols increasingly include early protein-rich enteral nutrition and, in some institutions, BCAA-enriched supplementation. Multiple randomized trials have shown that BCAA supplementation in the perioperative period reduces post-operative complications, accelerates wound healing, and shortens length of stay.
Burn recovery. Severe burn injury produces one of the most extreme hypermetabolic states in clinical medicine, with sustained protein breakdown that can persist for months and result in profound muscle wasting if not aggressively countered with high-protein nutrition. Modern burn care protocols include very high protein intake (2-3 g/kg/day, sometimes higher) with attention to BCAA delivery. Pediatric burn patients in particular benefit from intensive nutritional support including BCAAs.
Beyond these intensive-care contexts, more routine post-surgical and post-injury recovery also benefits from attention to dietary protein. Outpatient surgical patients (post-knee replacement, post-hysterectomy, post-cosmetic surgery, etc.) are often discharged with little nutritional guidance and may eat poorly during the early recovery period when their protein needs are elevated. Pre-operative dietary counseling (achieve and maintain adequate protein intake for 2-4 weeks before elective surgery, target 1.2-1.5 g/kg/day from varied protein sources) and post-operative continuation of this pattern produces better wound healing and faster functional recovery.
Chronic Wounds — Diabetic Ulcers and Pressure Sores
Chronic wounds — defined as wounds that fail to progress through the normal healing phases in the expected timeframe — are a major clinical and quality-of-life problem, particularly in older adults and patients with diabetes, peripheral vascular disease, or limited mobility. The most common chronic wounds are:
- Diabetic foot ulcers — multifactorial: peripheral neuropathy reduces protective sensation, peripheral arterial disease impairs perfusion, hyperglycemia impairs immune function, and chronic inflammation in the wound bed prevents transition from inflammatory to proliferative phase.
- Venous stasis ulcers — chronic venous insufficiency produces persistent edema, fibrin deposition around capillaries, and impaired wound oxygenation.
- Pressure injuries (decubitus ulcers, bed sores) — common in immobile patients (post-stroke, post-spinal-cord-injury, end-of-life care). Repeated mechanical compression of skin over bony prominences causes tissue ischemia and breakdown.
- Arterial ulcers — from peripheral arterial disease with critical limb ischemia. Limited blood flow severely restricts both substrate delivery and waste removal.
Patients with chronic wounds are frequently nutritionally compromised. They tend to be older, often have reduced appetite (sometimes related to the chronic inflammatory state of the wound itself), and may have multiple comorbidities affecting metabolism. Standard wound-care protocols increasingly include nutritional assessment and intervention. Adequate protein intake (1.2-1.5 g/kg/day, or higher in severely cachectic patients) with attention to BCAA and isoleucine adequacy is part of comprehensive wound management. Specific BCAA supplementation has been shown in some trials to improve wound healing rates in elderly patients with pressure ulcers.
The Mediterranean dietary pattern — rich in fish, legumes, nuts, eggs, dairy, and modest amounts of poultry — supplies adequate complete protein with isoleucine in physiologic balance, and is associated with better wound healing outcomes than less protein-rich Western dietary patterns. For wound-care patients without specific protein restrictions, encouraging Mediterranean-style eating with emphasis on protein at every meal is a reasonable nutritional foundation.
For diabetic wound care specifically, see our pages on Insulin Resistance and the related deep-dives. For the broader nutritional foundation of wound care, see Zinc (required for matrix metalloproteinase activity and epithelial proliferation) and Vitamin C (required for collagen hydroxylation).
Dietary Strategy for Hemoglobin and Healing Support
The practical dietary approach for ensuring adequate isoleucine to support hemoglobin synthesis and wound healing:
Foundation: 1.2-1.6 grams of total protein per kilogram body weight per day from varied sources. For an 80 kg adult, that is 96-128 grams of protein daily, distributed across 3-4 meals. This intake automatically supplies 3-6 grams of isoleucine per day — well above the WHO recommended minimum of 20 mg/kg/day (1.6 grams for an 80 kg adult).
Higher-need contexts (active recovery, post-surgery, chronic wound, severe illness, intense training, older adult sarcopenia prevention): target 1.6-2.0 grams of total protein per kilogram per day, with each meal containing at least 30 grams of complete protein.
Best food sources for combined hemoglobin and wound healing support:
- Beef and red meat — supplies isoleucine (and the full BCAA spectrum) alongside heme iron (the most bioavailable form for hemoglobin synthesis), zinc, B12, copper, and vitamin A precursors. The most efficient single source for hemoglobin support in mixed-diet eaters.
- Eggs — complete protein with excellent amino acid bioavailability, plus choline, B12, and selenium. Two large eggs supply approximately 0.7 grams of isoleucine and 12 grams of total protein.
- Salmon and other oily fish — complete protein with omega-3 fatty acids that support anti-inflammatory aspects of the healing response.
- Greek yogurt and cottage cheese — concentrated complete protein, convenient and shelf-stable, easy to consume in adequate quantity even when appetite is reduced.
- Whey protein — the highest BCAA content per gram of any practical protein source, useful for supplementing total intake without large meal volume.
- Lentils and soy — the plant sources with the highest isoleucine content per serving. Plant-based eaters can meet isoleucine needs with attention to including these regularly.
Combining complete protein sources at meals supports both the immediate amino acid availability and the long-term nutritional foundation for ongoing hemoglobin production and wound healing. For patients with established deficiency or active wound recovery, deliberate increase in protein-rich foods at every meal is the most reliable nutritional intervention.
For the broader context of dietary protein and amino acid needs, see the main Isoleucine page and the Muscle Protein Synthesis deep-dive.
Cautions
- Maple syrup urine disease (MSUD) — absolute contraindication. Patients with this rare metabolic disorder cannot catabolize BCAAs and require lifelong dietary restriction.
- Iron deficiency assessment first. If anemia is present, iron deficiency should be evaluated and ruled out or treated first. Iron deficiency is far more common than amino acid deficiency as a cause of anemia in developed-country populations.
- B12 and folate assessment in macrocytic anemia. The classic megaloblastic anemias are much more common than amino acid deficiency as a cause of macrocytic anemia.
- Renal disease — chronic kidney disease may require dietary protein restriction. Discuss protein targets with a renal dietitian before increasing intake for any of the reasons discussed on this page.
- Wound infection takes precedence over nutritional optimization. Active wound infection requires appropriate antimicrobial treatment, debridement, and drainage. Nutritional support is adjunctive, not primary.
- Beware oversimplification. Wound healing is multifactorial: tissue perfusion (oxygen delivery), absence of infection, glycemic control in diabetics, mechanical offloading of pressure points, zinc and vitamin C adequacy, smoking cessation, and many other factors. Isoleucine and protein adequacy are necessary but not sufficient for good wound healing outcomes.
- Vegan and vegetarian considerations — plant-based eaters with active wounds or anemia should be especially deliberate about combining protein sources. Pure fruitarian, raw food, or very restrictive macrobiotic diets are generally inappropriate during active recovery periods.
- Isolated BCAA supplements vs whole protein — whole protein is preferred for general hemoglobin and wound-healing support because it provides all the amino acids the body needs in physiologic balance. Isolated BCAA supplementation is a more specific intervention reserved for specific clinical situations (severe burns under intensivist supervision, hepatic encephalopathy under hepatologist supervision, post-surgical recovery in carefully selected patients).
Key Research Papers
- Perutz MF, Rossmann MG, Cullis AF, Muirhead H, Will G, North AC (1960). Structure of haemoglobin: a three-dimensional Fourier synthesis at 5.5-A resolution. Nature, 185(4711):416-422. — PubMed
- Schroeder WA et al. (1967). The amino acid sequence of the alpha-chain of bovine fetal haemoglobin. Biochimica et Biophysica Acta, 140(1):170-178. — PubMed
- Sanger F (1952). The arrangement of amino acids in proteins. Advances in Protein Chemistry, 7:1-67. — PubMed
- Sunkara LT, Achanta M, Schreiber NB et al. (2011). Butyrate enhances disease resistance of chickens by inducing antimicrobial host defense peptide gene expression. PLoS One, 6(11):e27225. — PubMed
- Mao X, Qi S, Yu B et al. (2013). Dietary L-arginine supplementation enhances porcine beta-defensins gene expression in some tissues of weaned pigs. Livestock Science, 154(1-3):167-170. — PubMed
- Sasaki E, Umeda T, Takahashi I et al. (2013). Effect of glutamine supplementation on neutrophil function in male judoists. Luminescence, 28(4):442-449. — PubMed
- Demling RH (2009). Nutrition, anabolism, and the wound healing process: an overview. Eplasty, 9:e9. — PubMed
- Stechmiller JK (2010). Understanding the role of nutrition and wound healing. Nutrition in Clinical Practice, 25(1):61-68. — PubMed
- Posthauer ME, Banks M, Dorner B, Schols JM (2015). The role of nutrition for pressure ulcer management: National Pressure Ulcer Advisory Panel, European Pressure Ulcer Advisory Panel, and Pan Pacific Pressure Injury Alliance white paper. Advances in Skin & Wound Care, 28(4):175-188. — PubMed
- Williams JZ, Abumrad N, Barbul A (2002). Effect of a specialized amino acid mixture on human collagen deposition. Annals of Surgery, 236(3):369-374. — PubMed
- De Bandt JP, Cynober L (2006). Therapeutic use of branched-chain amino acids in burn, trauma, and sepsis. Journal of Nutrition, 136(1 Suppl):308S-313S. — PubMed
- Wernerman J, Hammarqvist F, Vinnars E (1990). Alpha-ketoglutarate and postoperative muscle catabolism. The Lancet, 335(8691):701-703. — PubMed
- Bevins CL, Salzman NH (2011). Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nature Reviews Microbiology, 9(5):356-368. — PubMed
PubMed Topic Searches
- PubMed: Isoleucine hemoglobin globin synthesis
- PubMed: Bovine vs human hemoglobin sequence
- PubMed: BCAA beta-defensin epithelial
- PubMed: BCAA wound healing post-surgical
- PubMed: Protein-energy malnutrition anemia
Connections
- Isoleucine Overview
- Isoleucine Benefits Hub
- Isoleucine for Muscle Protein Synthesis
- Isoleucine for Glucose Uptake
- Isoleucine for Endurance Performance
- Leucine
- Valine
- Glycine (Heme Precursor)
- Anemia
- Iron
- Copper (Lysyl Oxidase)
- Zinc (Wound Healing)
- Vitamin C (Collagen)
- Organ Meats (Iron, B12)
- All Amino Acids