Isoleucine for Muscle Protein Synthesis
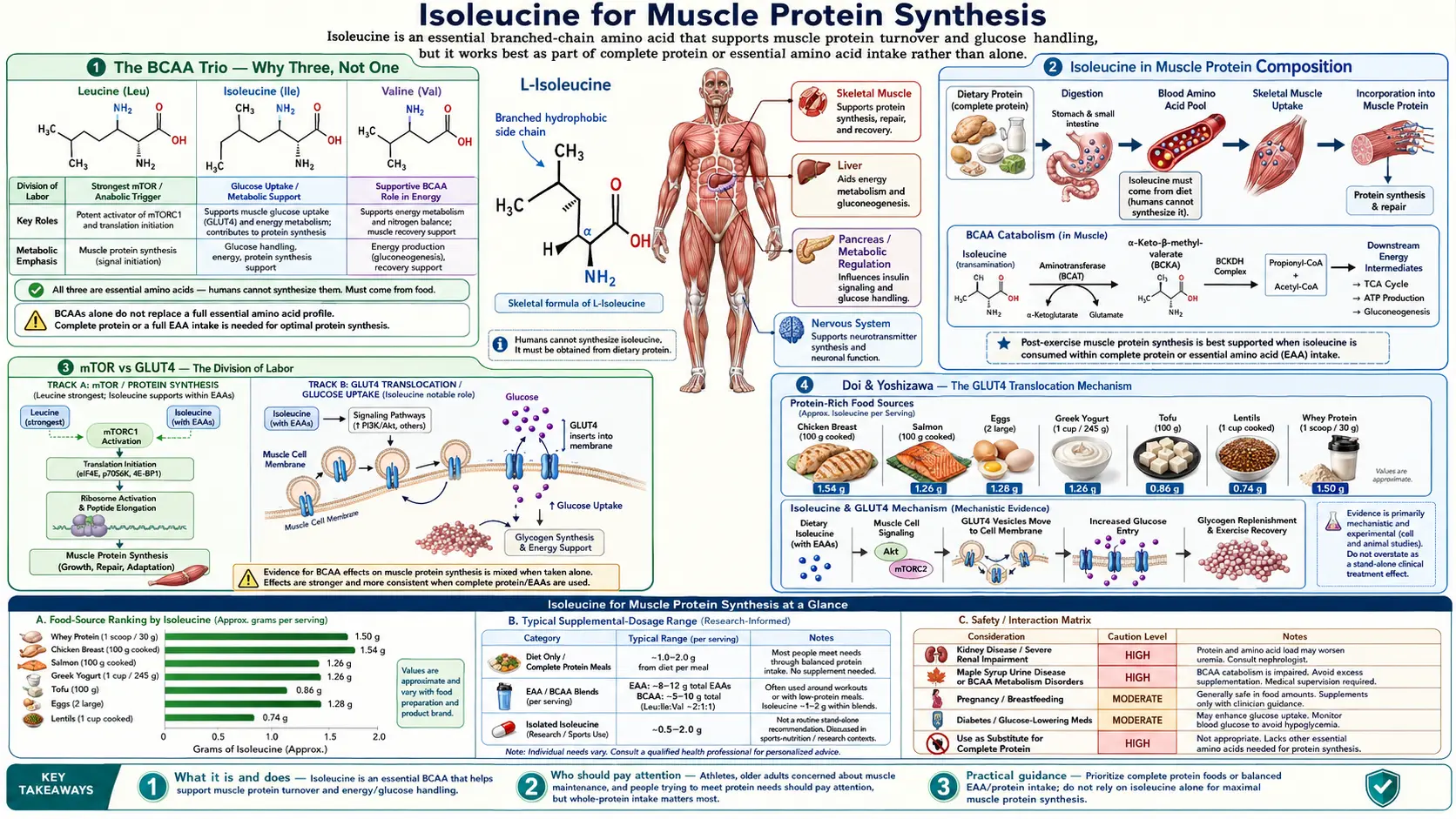
Isoleucine is one of three branched-chain amino acids (BCAAs) — alongside leucine and valine — that together comprise roughly 35% of the essential amino acids in human muscle protein. While leucine has captured most of the popular attention as the dominant mTORC1 activator, isoleucine plays a structurally distinct and complementary role: it is the BCAA most strongly associated with insulin-independent GLUT4 translocation and glucose delivery to working muscle, the substrate stream that fuels the actual peptide-bond formation step. The canonical 2:1:1 leucine:isoleucine:valine ratio found in nearly every commercial BCAA supplement is not arbitrary — it reflects both the proportion in mammalian skeletal muscle and the empirical ratio that produces optimal recovery in human exercise trials. This deep-dive walks through the molecular mechanism of isoleucine's role in muscle anabolism, how it differs from leucine's mTOR effect, and what the evidence actually says about isolated isoleucine versus full-spectrum BCAA dosing.
Table of Contents
- The BCAA Trio — Why Three, Not One
- Isoleucine in Muscle Protein Composition
- mTOR vs GLUT4 — The Division of Labor
- Doi & Yoshizawa — The GLUT4 Translocation Mechanism
- The 2:1:1 Leucine:Isoleucine:Valine Ratio
- Anti-Catabolic Effect During Caloric Restriction
- Exercise Timing — Pre, Intra, and Post
- Sarcopenia, Anabolic Resistance, and Older Adults
- Evidence Quality — What the Trials Actually Show
- Cautions and Limitations
- Key Research Papers
- Connections
- Featured Videos
The BCAA Trio — Why Three, Not One
Leucine, isoleucine, and valine are grouped together as the branched-chain amino acids (BCAAs) because of a shared structural feature: each has an aliphatic side chain with a branch (a methyl group projecting off the main carbon backbone). This branching is more than a chemistry trivia point — it dictates how the three amino acids are metabolized. Unlike most amino acids, which are deaminated and processed primarily in the liver, the BCAAs largely bypass the liver and undergo their first metabolic step (transamination by the BCAA aminotransferase, BCAT) in skeletal muscle, kidney, and brain.
The shared metabolic pathway is biologically significant for three reasons. First, the BCAAs use the same enzymatic machinery for catabolism: the branched-chain alpha-keto acid dehydrogenase (BCKDH) complex processes the alpha-keto acids derived from all three. A genetic defect in BCKDH causes maple syrup urine disease (MSUD), which simultaneously affects leucine, isoleucine, and valine metabolism. Second, the BCAAs compete with one another and with the aromatic amino acids (tryptophan, tyrosine, phenylalanine) for transport into cells and across the blood-brain barrier via the L-type amino acid transporter (LAT1). Third, because their fates are linked, supplementing any one BCAA in isolation can perturb the balance and produce unexpected effects on the other two.
Despite the shared pathway, each BCAA has distinct downstream effects. Leucine is the dominant activator of the mechanistic target of rapamycin complex 1 (mTORC1), the master regulator of protein synthesis. Isoleucine has the strongest effect on glucose uptake and energy substrate delivery to muscle. Valine's role is the least well-characterized of the three but appears to involve energy metabolism in the brain and modulation of central fatigue. The three working together produce the integrated anabolic response that no single BCAA produces in isolation — which is why the canonical supplement ratio includes all three rather than just leucine.
Isoleucine in Muscle Protein Composition
Human skeletal muscle protein is approximately 35% BCAAs by weight of essential amino acids, with isoleucine accounting for roughly 8-9% of the total amino acid content of myofibrillar protein. This is a higher concentration than most non-muscle tissues, which is one reason why post-exercise muscle damage produces a measurable efflux of BCAAs into the bloodstream — the damaged tissue releases what it was disproportionately built from.
The structural role of isoleucine within muscle proteins is concentrated in the hydrophobic interior of folded protein domains. The isoleucine side chain (sec-butyl group) is one of the most hydrophobic among the 20 standard amino acids, and it preferentially partitions into the water-excluding core of globular protein structures. Actin, myosin heavy chain, troponin, tropomyosin, and titin all contain isoleucine residues at functionally critical positions where the side chain's hydrophobic character stabilizes the fold or the interaction with another protein.
A practical consequence: when muscle protein is broken down and rebuilt (the constant turnover that occurs even at rest, plus the accelerated turnover that follows exercise), the new protein needs isoleucine in the same proportion. The post-translational fidelity of muscle protein synthesis depends on a free pool of isoleucine in the cell that can be loaded onto its dedicated tRNA (tRNA-Ile) by isoleucyl-tRNA synthetase. If isoleucine availability drops below a threshold, ribosomes stall at isoleucine codons, the integrated stress response is triggered, and global protein synthesis is downregulated even when leucine and other amino acids remain abundant.
mTOR vs GLUT4 — The Division of Labor
The cleanest way to understand the distinct roles of leucine and isoleucine in muscle anabolism is to think of them as operating in parallel on two different limiting steps of protein synthesis: signal versus substrate.
Leucine provides the signal. When intracellular leucine concentration rises above a threshold (the leucine sensor is a protein called sestrin2, which binds leucine and releases its inhibition of mTORC1), the mTORC1 kinase complex is activated. mTORC1 then phosphorylates downstream targets including p70 S6 kinase and 4E-BP1, which together upregulate translation initiation and elongation. The net effect is that the ribosomes "turn on" protein synthesis.
Isoleucine helps deliver the substrate. Independent of mTOR, isoleucine activates the PI3K/Akt signaling cascade, which results in translocation of GLUT4 glucose transporters from intracellular storage vesicles to the muscle cell membrane. Once at the surface, GLUT4 allows glucose to enter the cell down its concentration gradient. The imported glucose is used both to replenish muscle glycogen (the energy reserve) and to provide carbon skeletons for non-essential amino acid synthesis. This is the substrate stream that the ribosomes need once leucine has turned them on.
The division of labor is not absolute — leucine also has some effect on glucose uptake, and isoleucine contributes weakly to mTORC1 activation — but the proportional emphasis is distinct. A muscle cell exposed to leucine alone has ribosomes ready to translate but limited substrate to translate with. A muscle cell exposed to isoleucine alone has plenty of substrate but ribosomes that are not maximally activated. The two together produce a multiplicative effect that exceeds the sum of either alone.
This is mechanistically why leucine-only supplementation (occasionally marketed as "the only BCAA you need") tends to underperform full-spectrum BCAA supplementation in human trials. Leucine without isoleucine and valine can even paradoxically reduce circulating isoleucine and valine concentrations because the activated mTORC1 pathway accelerates uptake of all three into muscle, depleting the plasma pool of the two that were not supplemented.
Doi & Yoshizawa — The GLUT4 Translocation Mechanism
The clearest molecular characterization of isoleucine's glucose-uptake effect came from a series of studies by Masako Doi, Asami Yoshizawa, and colleagues at the Ajinomoto Institute of Life Sciences in Japan in the early 2000s. Their 2003 paper in Biochemical and Biophysical Research Communications demonstrated that isoleucine, administered alone to fasted rats, lowered plasma glucose concentration through a mechanism that did not require insulin. Pancreatectomized rats (no endogenous insulin) and rats pretreated with anti-insulin antibodies still showed the isoleucine-induced glucose-lowering effect, confirming the insulin-independence.
Follow-up work in 2005 mapped the molecular mechanism. Isoleucine administration activated PI3-kinase in skeletal muscle (measured by phosphoinositide phosphorylation assays) and triggered GLUT4 translocation from intracellular vesicles to the sarcolemma (measured by membrane fractionation and Western blot). Critically, the effect on PI3-kinase was independent of insulin receptor activation — isoleucine engaged a parallel signaling input upstream of PI3K. The downstream phosphorylation of Akt and the resulting GLUT4 translocation were quantitatively similar to those produced by a moderate insulin dose, but reached through a different upstream route.
The clinical significance: in conditions of insulin resistance — type 2 diabetes, metabolic syndrome, obesity — the insulin signaling cascade is impaired at the insulin receptor or insulin receptor substrate level. Isoleucine's parallel pathway is not subject to the same defect, which means it can drive glucose uptake into muscle in patients whose insulin signaling has degraded. This is the mechanistic basis for the proposed use of isoleucine as a metabolic support nutrient in insulin-resistant states, although clinical trial evidence in humans is still developing and the picture is complicated by the BCAA paradox (see the Glucose Uptake deep-dive for the full discussion).
From a muscle protein synthesis perspective, the Doi findings explain why isoleucine's role in muscle anabolism is not redundant with leucine's. The two operate on different steps. Leucine activates the synthetic machinery. Isoleucine ensures that the substrate (glucose, which provides both energy and carbon for non-essential amino acid synthesis) actually reaches the inside of the cell where it can be used.
The 2:1:1 Leucine:Isoleucine:Valine Ratio
The standard BCAA supplement ratio is 2 parts leucine to 1 part isoleucine to 1 part valine, by mass. This ratio is not a marketing convention — it is grounded in three converging lines of evidence:
- Skeletal muscle composition. Direct amino acid analysis of human skeletal muscle protein shows that the leucine:isoleucine:valine ratio in muscle is approximately 2:1:1. If the goal of BCAA supplementation is to rebuild muscle, providing the constituent amino acids in their natural proportions is a reasonable first-pass strategy.
- Dose-response in mTORC1 activation. Leucine has a higher EC50 (the concentration required for half-maximal effect) on mTORC1 than isoleucine or valine, meaning more leucine is needed per unit signaling output. Double the leucine partially compensates for this differential potency.
- Empirical trial data. Multiple human exercise studies comparing 2:1:1 to 4:1:1, 8:1:1, and leucine-only formulations have found that 2:1:1 produces the most consistent recovery and muscle protein synthesis outcomes. Higher leucine ratios do not produce additional benefit and may impair isoleucine and valine availability.
The 2:1:1 ratio also corresponds approximately to the natural ratio in high-quality animal protein sources. Whey protein, for example, has roughly 12% leucine, 6% isoleucine, and 6% valine by amino acid mass — the 2:1:1 ratio is essentially the ratio of these three in whey. Eating whey or other complete protein delivers the three BCAAs in physiologically appropriate proportions automatically.
Some specialized formulations deviate from 2:1:1 for specific purposes. Hepatic encephalopathy BCAA formulas use a higher BCAA:aromatic ratio (the goal is to displace aromatic amino acids from the brain, not to optimize muscle anabolism). Endurance-focused formulas sometimes use higher valine to support central fatigue resistance (see the Endurance Performance deep-dive). For general muscle anabolic support, 2:1:1 remains the well-validated default.
Anti-Catabolic Effect During Caloric Restriction
Beyond stimulating protein synthesis, isoleucine and the other BCAAs help blunt protein breakdown during catabolic conditions. The mechanism operates through two parallel routes: substrate competition and signaling.
In substrate terms, when the body is in negative energy balance (caloric restriction, fasting, prolonged endurance exercise), it begins breaking down muscle protein to liberate amino acids for use as gluconeogenic substrates and energy. The branched-chain amino acids can be directly oxidized in muscle to provide energy without requiring hepatic processing first. If BCAAs are available exogenously (from supplementation or recent food intake), the muscle preferentially oxidizes the dietary BCAAs rather than catabolizing its own structural proteins to obtain them. This is a sparing effect: the muscle's own protein is protected because the alternative fuel is available.
In signaling terms, leucine acts through mTORC1 to inhibit autophagy — the process by which cells degrade their own components (including muscle proteins) to recycle the amino acids and salvage energy. Activated mTORC1 phosphorylates ULK1, blocking initiation of autophagy. Isoleucine contributes to this anti-autophagic effect indirectly through its support of substrate availability that keeps the cell in an anabolic state.
The clinical applications include bariatric surgery patients (very large protein deficit during the post-operative weight loss phase, with high risk of lean mass loss), cancer cachexia patients (chronic inflammatory catabolism), critical illness (ICU-related muscle wasting), and the elderly on calorie-restricted diets. In each case, ensuring adequate BCAA intake — either through deliberate inclusion of leucine/isoleucine/valine-rich protein at meals or through targeted supplementation — helps preserve lean mass during the period of negative energy balance.
For athletes doing deliberate cutting phases (intentional caloric restriction to reduce body fat while preserving muscle), BCAA supplementation between meals is a common strategy. The most-cited supplementation pattern is 5-10 grams of 2:1:1 BCAAs taken 30 minutes before fasted training, with additional dosing at mid-day if there is a long gap between meals. The goal is not to add calories — BCAA supplements provide minimal energy — but to maintain the anti-catabolic signaling during periods when whole-protein intake is intentionally reduced.
Exercise Timing — Pre, Intra, and Post
Three timing windows around exercise have been studied for BCAA supplementation, each with a distinct rationale and supporting evidence base:
Pre-exercise (15-30 minutes before). Loading BCAAs before training puts isoleucine into the bloodstream during the workout, where it can support GLUT4-mediated glucose uptake into working muscle. This is most relevant for fasted training, where there is no recent protein meal to provide the same effect. Typical dose: 5-10 grams of 2:1:1 BCAAs in water. For fed athletes who eat a normal meal 2-3 hours before training, the pre-workout BCAA dose adds little marginal benefit.
Intra-exercise (during long workouts). For endurance sessions exceeding 90 minutes, BCAA consumption during the workout can support central fatigue resistance (by limiting tryptophan uptake into the brain — the Endurance Performance deep-dive explores this in detail) and provide oxidative substrate to working muscle. Typical dose: 5-7 grams of 2:1:1 BCAAs in a flavored sports drink consumed in small sips throughout the session.
Post-exercise (immediately to 60 minutes after). The traditional "anabolic window" for protein consumption. The evidence here is the most robust: a meal or supplement containing 20-40 grams of high-quality protein (which inherently includes 4-8 grams of BCAAs) within an hour of resistance training maximizes muscle protein synthesis. Isolated BCAA supplementation in this window has been shown to elevate muscle protein synthesis, but the effect is consistently smaller than that of a complete protein source containing the full essential amino acid spectrum. The current consensus is that whole protein (whey, egg, casein, mixed-source) is preferred post-workout, with isolated BCAAs reserved for fasted training, between-meal anti-catabolic protection, or athletes who cannot tolerate a full protein meal immediately after exercise.
For the typical resistance-training enthusiast eating 1.6-2.2 grams of protein per kilogram of body weight per day from varied sources, the marginal benefit of isolated BCAA supplementation is small. The BCAA market is driven more by the convenience and flavor of liquid intra-workout drinks than by clearly demonstrated incremental physiological benefit over a balanced protein intake.
Sarcopenia, Anabolic Resistance, and Older Adults
Sarcopenia — the age-related loss of skeletal muscle mass and function — is a primary driver of functional decline, falls, fractures, and loss of independence in older adults. The mechanism is multifactorial, but a central component is anabolic resistance: the diminished muscle protein synthetic response to a given dose of dietary protein in older adults compared to younger adults.
Older adults need a higher leucine threshold (estimated at 2.5-3.0 grams of leucine per meal, versus 1.5-2.0 grams in younger adults) to maximally trigger mTORC1 activation and post-prandial muscle protein synthesis. This threshold corresponds to approximately 30-40 grams of high-quality protein per meal, distributed across three or four meals per day. Many older adults fall well short of this target, particularly at breakfast, which tends to be the lowest-protein meal of the day.
Isoleucine's role in this context is twofold. First, the GLUT4-translocation mechanism may help compensate for the age-related decline in insulin sensitivity. Older muscle is less responsive to insulin signaling, and the alternative isoleucine-driven pathway provides a parallel route for substrate delivery. Second, the anti-catabolic effect of BCAAs is particularly relevant in older adults, who often experience accelerated muscle protein breakdown during periods of bed rest, hospitalization, or acute illness — the periods when sarcopenia tends to accelerate dramatically.
Practical recommendations for older adults at risk for sarcopenia include: distribute protein evenly across meals (rather than back-loading at dinner), prioritize complete protein sources at each meal (whey, eggs, meat, fish, dairy, or well-combined plant proteins), pair protein intake with resistance training when medically appropriate, and consider leucine-enriched BCAA supplementation (40% leucine formulations exist for this purpose) during recovery from illness or surgery. Whole protein remains the first choice, with isolated BCAAs as an adjunct when oral intake is reduced.
For the broader management of muscle wasting, see our pages on Fatigue and the general Leucine deep-dive.
Evidence Quality — What the Trials Actually Show
BCAA supplementation has been studied in hundreds of randomized controlled trials, with widely varying methodology, populations, and outcomes. A frank summary of where the evidence is strong versus weak:
Robust evidence: BCAA supplementation reduces exercise-induced muscle soreness (DOMS) modestly, accelerates recovery of muscle function after eccentric exercise, and reduces markers of muscle damage (creatine kinase, lactate dehydrogenase) post-exercise. The effect sizes are typically small to moderate — helpful but not transformative.
Mixed evidence: Isolated BCAA supplementation versus a complete protein source for post-exercise muscle protein synthesis. Most direct head-to-head trials favor complete protein. BCAAs are not a replacement for adequate dietary protein; they are at most a supplement to it.
Weak or contradictory evidence: BCAA supplementation as a fat-loss aid (claimed mechanism: BCAAs spare muscle while accelerating fat oxidation; evidence: minimal). BCAA supplementation for general athletic performance enhancement in athletes who already consume adequate dietary protein. Isolated isoleucine supplementation as a diabetes treatment (the mechanistic basis is plausible, but clinical trials in humans have produced inconsistent results, and the BCAA paradox raises concerns about long-term high-dose supplementation).
The honest summary for the typical reader: if your dietary protein intake is adequate (roughly 1.2-1.6 grams per kilogram body weight per day from varied sources, with at least 25-40 grams of complete protein per meal), you are getting plenty of isoleucine and BCAAs from food. Adding a BCAA supplement on top of an already-adequate diet produces small or no additional benefit for most outcomes. If your dietary protein intake is inadequate (vegan athletes who do not consume enough varied plant protein, older adults with low appetite, patients with malabsorption), BCAA supplementation can fill specific gaps but is generally not as effective as simply consuming more complete protein.
The narrowest reasonable use case for isolated BCAAs is fasted training (where pre-workout BCAAs offer anti-catabolic protection without breaking the fast in any meaningful caloric sense) and acute clinical conditions (hepatic encephalopathy, certain ICU protocols) where the specific BCAA ratio matters more than the calorie or complete-protein delivery.
Cautions and Limitations
- Maple syrup urine disease (MSUD) — the absolute contraindication. Patients with MSUD have defective BCKDH and cannot metabolize BCAAs. Elevated plasma BCAAs are neurotoxic. Standard MSUD dietary management severely restricts isoleucine, leucine, and valine to the minimum required for protein synthesis. Any BCAA supplementation in an MSUD patient is potentially life-threatening.
- Advanced liver disease (hepatic encephalopathy) — the relationship is complex. Standard BCAAs reduce ammonia and improve encephalopathy symptoms in cirrhosis. However, the dosing and form (specialized hepatic-encephalopathy BCAA formulas, not over-the-counter products) should be supervised by a hepatologist.
- Type 2 diabetes and insulin resistance — the BCAA paradox. Cross-sectional studies consistently show elevated serum BCAA concentrations in insulin-resistant individuals, and the elevation predicts subsequent diabetes risk. Whether chronic BCAA supplementation worsens or improves the underlying insulin resistance remains debated. Patients with established T2D should not self-supplement high-dose BCAAs without physician oversight.
- Pregnancy and lactation — adequate isoleucine from a balanced protein-rich diet is required for fetal growth and milk production. Isolated BCAA supplementation in pregnancy is not well-studied and is generally not recommended. Whole protein from food is preferred.
- Renal impairment — advanced chronic kidney disease requires dietary protein restriction. Adding BCAA supplements increases nitrogen load and can accelerate uremic symptoms. Discuss with a renal dietitian before supplementing.
- The leucine ceiling — high-dose isolated leucine (above 5-10 grams in a single dose) can depress plasma isoleucine and valine via shared transport competition and accelerated BCKDH catabolism. Leucine-only supplementation paradoxically lowers the partner BCAAs and is not recommended outside specific research contexts.
- GI tolerance — high-dose BCAAs (above 20 grams in a single serving) commonly produce GI upset, nausea, and cramping. Split-dosing across the day is better tolerated.
Key Research Papers
- Doi M, Yamaoka I, Fukunaga T, Nakayama M (2003). Isoleucine, a potent plasma glucose-lowering amino acid, stimulates glucose uptake in C2C12 myotubes. Biochemical and Biophysical Research Communications, 312(4):1111-1117. — PubMed
- Doi M, Yamaoka I, Nakayama M, Sugahara K, Yoshizawa F (2005). Hypoglycemic effect of isoleucine involves increased muscle glucose uptake and whole body glucose oxidation and decreased hepatic gluconeogenesis. American Journal of Physiology - Endocrinology and Metabolism, 292(6):E1683-1693. — PubMed
- Anthony JC et al. (2000). Orally administered leucine stimulates protein synthesis in skeletal muscle of postabsorptive rats. Journal of Nutrition, 130(2):139-145. — PubMed
- Wolfe RR (2017). Branched-chain amino acids and muscle protein synthesis in humans: myth or reality? Journal of the International Society of Sports Nutrition, 14:30. — PubMed
- Jackman SR et al. (2017). Branched-chain amino acid ingestion stimulates muscle myofibrillar protein synthesis following resistance exercise in humans. Frontiers in Physiology, 8:390. — PubMed
- Plotkin DL et al. (2021). Isolated leucine and branched-chain amino acid supplementation for enhancing muscular strength and hypertrophy: a narrative review. International Journal of Sport Nutrition and Exercise Metabolism, 31(3):292-301. — PubMed
- Wolfson RL et al. (2016). Sestrin2 is a leucine sensor for the mTORC1 pathway. Science, 351(6268):43-48. — PubMed
- Phillips SM (2014). A brief review of critical processes in exercise-induced muscular hypertrophy. Sports Medicine, 44 Suppl 1:S71-77. — PubMed
- Kimball SR, Jefferson LS (2006). Signaling pathways and molecular mechanisms through which branched-chain amino acids mediate translational control of protein synthesis. Journal of Nutrition, 136(1 Suppl):227S-231S. — PubMed
- Churchward-Venne TA et al. (2014). Leucine supplementation of a low-protein mixed macronutrient beverage enhances myofibrillar protein synthesis in young men: a double-blind, randomized trial. American Journal of Clinical Nutrition, 99(2):276-286. — PubMed
- Bauer J et al. (2013). Evidence-based recommendations for optimal dietary protein intake in older people: a position paper from the PROT-AGE Study Group. JAMDA, 14(8):542-559. — PubMed
- Layman DK (2003). The role of leucine in weight loss diets and glucose homeostasis. Journal of Nutrition, 133(1):261S-267S. — PubMed
PubMed Topic Searches
- PubMed: Isoleucine muscle protein synthesis mTOR
- PubMed: 2:1:1 BCAA ratio
- PubMed: Isoleucine GLUT4 translocation PI3K
- PubMed: BCAA anti-catabolic muscle sparing
- PubMed: Sarcopenia leucine threshold anabolic resistance
Connections
- Isoleucine Overview
- Isoleucine Benefits Hub
- Isoleucine for Glucose Uptake
- Isoleucine for Endurance Performance
- Isoleucine for Hemoglobin & Wound Healing
- Leucine
- Valine
- Glutamine
- Arginine
- All Amino Acids
- Insulin Resistance
- Metabolic Syndrome
- Fatigue
- Eggs