Histidine, Carnosine, and Antioxidant Defense
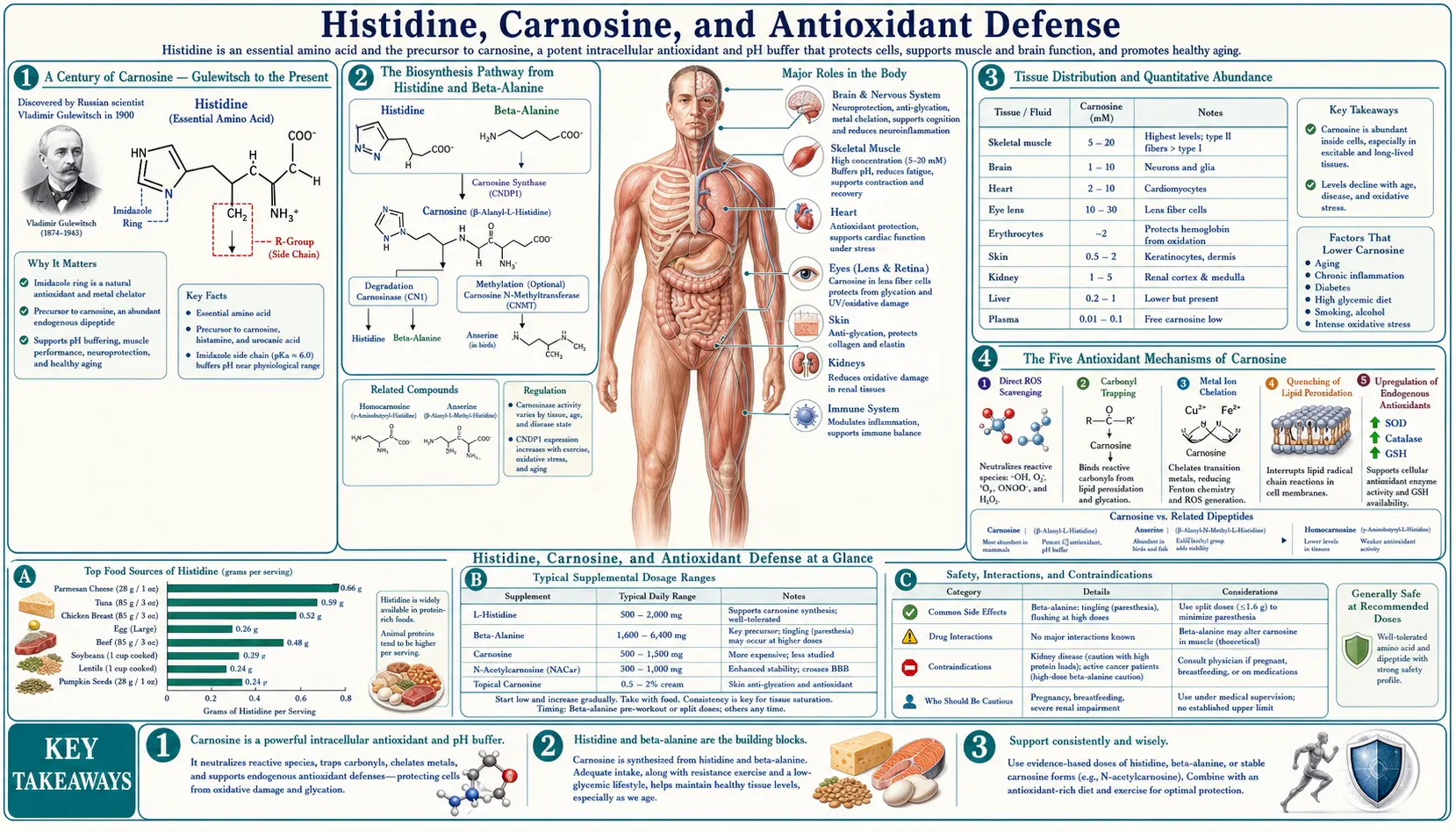
Carnosine is the second-most-abundant nitrogen compound in muscle tissue (after creatine), and the body manufactures essentially all of it from histidine and beta-alanine using a single ATP-dependent ligase. Russian biochemist V. S. Gulewitsch first isolated carnosine from a Liebig's meat extract in 1900 and named it after the Latin carnis (flesh) — the very word "carnivore" shares the same root. Over the next century, carnosine emerged as one of the most multi-functional small molecules in biology: a pH buffer (most of muscle's lactate-tolerance comes from carnosine), a direct free-radical scavenger, a specific quencher of the lipid peroxidation product 4-hydroxynonenal, a chelator of pro-oxidant copper and iron, and the most efficient endogenous inhibitor of advanced glycation end-product (AGE) formation. Muscle carnosine concentration declines roughly 60% between ages 25 and 70, mirroring the broader age-related rise in oxidative stress and protein damage. The dipeptide also exists as a related compound anserine (beta-alanyl-N-methylhistidine) in poultry and fish muscle — chicken breast and tuna are nature's richest carnosine/anserine sources. This deep-dive walks through the carnosine biosynthesis pathway from histidine and beta-alanine, the five distinct antioxidant mechanisms, the AGE-quenching role in diabetes complications, the age-related decline, and the practical question of whether to supplement histidine, beta-alanine (the rate-limiting precursor), or carnosine itself.
Table of Contents
- A Century of Carnosine — Gulewitsch to the Present
- The Biosynthesis Pathway from Histidine and Beta-Alanine
- Tissue Distribution and Quantitative Abundance
- The Five Antioxidant Mechanisms of Carnosine
- Advanced Glycation End-Product (AGE) Quenching
- Carnosine in Diabetic Complications
- Age-Related Decline of Muscle Carnosine
- Anserine in Poultry and Fish, Balenine in Whale Muscle
- Brain Carnosine and Polaprezinc (Zinc-Carnosine)
- Supplementation: Histidine, Beta-Alanine, or Carnosine?
- Key Research Papers
- Connections
- Featured Videos
A Century of Carnosine — Gulewitsch to the Present
Carnosine was first isolated in 1900 by Vladimir Sergeyevich Gulewitsch at the University of Moscow from a Liebig's meat extract — a 19th-century beef bouillon concentrate that was a staple convenience food and laboratory reagent of the era. Gulewitsch named the compound carnosine, from the Latin carnis (flesh), reflecting both its source and its abundance in muscle. The structure as a beta-alanyl-histidine dipeptide was determined a decade later. The closely related compound anserine (beta-alanyl-N-methylhistidine) was isolated from goose muscle in 1929 — the name comes from anser, Latin for goose. A third relative, balenine (beta-alanyl-N-tau-methylhistidine), was identified in whale muscle in 1962.
For most of the 20th century, carnosine was treated as a curiosity — abundant in muscle, but with no clear function beyond a vague role in pH buffering. The renaissance came in the 1980s and 1990s through the work of Russian biochemist Alexander A. Boldyrev and his colleagues at Moscow State University, who systematically demonstrated carnosine's direct antioxidant activity, its quenching of advanced glycation end-products, its protective effects in cardiac ischemia, and its specific role in scavenging the lipid peroxidation product 4-hydroxynonenal. By the early 2000s, carnosine had been recognized as one of the most multi-functional small molecules in muscle biology, and the broader implications — for diabetes complications, neurodegenerative disease, age-related sarcopenia, and exercise performance — began to emerge in the Western literature.
The exercise-physiology angle drove the commercial supplement industry. Roger Harris and Craig Sale (UK) and a number of Asian groups demonstrated in the early 2000s that beta-alanine supplementation (the rate-limiting precursor) dose-dependently raises muscle carnosine, and that the elevated muscle carnosine measurably improves high-intensity exercise performance by increasing intramuscular buffering capacity. Beta-alanine became one of the few sports supplements with strong RCT evidence for ergogenic effect, and the "tingling" sensation from beta-alanine ingestion (paresthesia from MRGPRD receptor activation) became a recognizable feature of pre-workout supplement formulations.
The Biosynthesis Pathway from Histidine and Beta-Alanine
Carnosine biosynthesis is remarkably simple compared to most peptide synthesis pathways. The body does not assemble carnosine on the ribosome (which would require a tRNA for beta-alanine, which does not exist). Instead, a dedicated soluble enzyme, carnosine synthase (ATPGD1, encoded by the CARNS1 gene on chromosome 11p15.4), directly ligates beta-alanine to L-histidine, using ATP hydrolysis to drive the reaction:
beta-alanine + L-histidine + ATP → carnosine + AMP + PPi
The enzyme has a Km for histidine of approximately 16 micromolar (well below muscle histidine concentration of around 300 micromolar) and a Km for beta-alanine of approximately 2 millimolar (around the muscle beta-alanine concentration). This kinetic asymmetry is critical — histidine is essentially always saturating, while beta-alanine is essentially always rate-limiting for carnosine synthesis. The implication: you cannot raise muscle carnosine by supplementing histidine alone (you are already saturating the enzyme on the histidine side), but you can substantially raise it by supplementing beta-alanine (which is below the Km and therefore directly controls the synthesis rate).
Carnosine synthase is expressed at highest levels in skeletal muscle, with substantial expression also in cardiac muscle, kidney, and certain brain regions (olfactory bulb, ventral mesencephalon). The enzyme is essentially absent from liver and from erythrocytes, which is why circulating plasma carnosine is very low (most of it is rapidly degraded by serum carnosinase) and almost all of the body's carnosine is located inside the cells where it was synthesized.
The serum carnosinase enzyme (CN1, encoded by CNDP1 on chromosome 18q22) is a critical regulator of carnosine pharmacokinetics. CN1 degrades free carnosine in plasma back to histidine and beta-alanine within minutes. This is why oral carnosine supplementation is largely ineffective for raising muscle carnosine in adults — the carnosine is degraded in the gut and bloodstream before reaching muscle. Beta-alanine supplementation works because beta-alanine is resistant to CN1 and reaches muscle in intact form for in-situ synthesis.
CNDP1 genetic variants have clinical relevance. A trinucleotide repeat polymorphism in CNDP1 (the "Mannheim allele") produces a less-active CN1 enzyme. Carriers have higher plasma carnosine and reduced risk of diabetic nephropathy in type 2 diabetes — one of the strongest genetic protective signals against diabetic kidney disease ever identified.
Tissue Distribution and Quantitative Abundance
Carnosine is one of the most abundant nitrogen compounds in skeletal muscle, second only to creatine and creatine phosphate. Type II (fast-twitch, glycolytic) muscle fibers contain approximately twice as much carnosine as type I (slow-twitch, oxidative) fibers, reflecting the greater need for pH buffering during anaerobic glycolysis.
Typical concentrations:
- Human vastus lateralis (mixed-fiber thigh muscle): 15-30 mmol/kg dry muscle
- Type II human muscle fibers (after biopsy fiber typing): 30-40 mmol/kg dry
- Type I human muscle fibers: 12-18 mmol/kg dry
- Sprinter / power athlete muscle: can reach 35-45 mmol/kg dry, reflecting both fiber-type composition and training adaptation
- Cardiac muscle: 5-10 mmol/kg dry
- Olfactory bulb (the highest-carnosine brain region): 1-3 mmol/kg wet
- Most other brain regions: below 0.5 mmol/kg wet
For comparison, ATP in muscle is around 5-8 mmol/kg dry, creatine plus phosphocreatine is around 50-80 mmol/kg dry. Carnosine is therefore in the same molar range as ATP and accounts for a sizable fraction of total muscle nitrogen.
The sex difference is real but modest — men typically have 20-30% higher muscle carnosine than women at the same training status, consistent with the higher type II fiber proportion in male muscle and possibly with androgen-driven effects on beta-alanine handling.
The dietary contribution to muscle carnosine is substantial in mixed omnivorous diets. Each 100g of beef contains approximately 365 mg of carnosine; chicken breast approximately 750 mg; pork around 425 mg. A typical Western omnivore intake of 100-300 mg of carnosine per day is rapidly degraded by serum carnosinase to beta-alanine and histidine, which then re-enter the muscle for in-situ resynthesis. Strict vegetarians and vegans have measurably lower muscle carnosine (typically 20-40% lower) than omnivores, reflecting the absence of dietary beta-alanine and dietary carnosine. This is one of the few clearly-demonstrated nutrient deficits of plant-based eating that is not easily replaced by plant food sources (beta-alanine is almost absent from plant foods).
The Five Antioxidant Mechanisms of Carnosine
Carnosine's antioxidant activity is multifactorial, with five distinct mechanisms that together make it more than just another general-purpose radical scavenger:
1. Direct hydroxyl radical scavenging. The imidazole ring (contributed by the histidine moiety) reacts with hydroxyl radicals at near-diffusion-limited rate constants. This is the same chemistry that makes free histidine an antioxidant, but the dipeptide is more stable and has a longer intracellular half-life than free histidine, providing more sustained protection.
2. Singlet oxygen quenching. Carnosine quenches singlet oxygen (a high-energy excited form of O2 generated by photochemistry and certain enzyme reactions) with a rate constant approximately 10-fold higher than histidine and 100-fold higher than glutathione. This is particularly relevant in tissues exposed to light (eye, skin) and in tissues with high peroxidase activity.
3. Specific quenching of 4-hydroxynonenal (4-HNE). The most distinctive carnosine activity. 4-HNE is a particularly toxic lipid peroxidation product — an alpha-beta unsaturated aldehyde that forms when polyunsaturated fatty acids undergo radical-mediated peroxidation. 4-HNE forms covalent adducts with lysine, cysteine, and histidine residues in proteins, damaging enzymes and triggering pro-inflammatory cell signaling. Carnosine reacts with 4-HNE via a Michael addition between the imidazole ring of the histidine moiety and the alpha-beta unsaturated carbonyl of 4-HNE, forming a stable carnosine-4-HNE adduct that is then excreted. This effectively detoxifies 4-HNE before it can damage essential proteins. The reaction is approximately 100-fold faster than the corresponding reaction with free histidine, reflecting kinetic catalysis from the beta-alanine moiety and from the imidazole orientation in the dipeptide.
4. Transition metal chelation. Carnosine chelates copper (II) and zinc with moderate affinity, and iron (II) more weakly. This is biologically significant because both free copper and free iron catalyze the Fenton reaction (Fe2+ or Cu+ + H2O2 → OH· + OH- + Fe3+/Cu2+), which generates hydroxyl radicals from cellular hydrogen peroxide. By sequestering catalytic metals, carnosine reduces the rate of hydroxyl radical formation in tissues with metal overload or under oxidative stress.
5. Inhibition of advanced glycation end-product (AGE) formation. The largest and most clinically important carnosine activity. Discussed in detail in the next section.
These five mechanisms together explain why carnosine activity is hard to reproduce with simpler general antioxidants — vitamin C, vitamin E, and glutathione each cover a subset of these activities, but no single small molecule covers all five. Carnosine is unique in this respect, which is why its decline with age has such a noticeable systemic effect.
Advanced Glycation End-Product (AGE) Quenching
Advanced glycation end-products (AGEs) are formed when reducing sugars (primarily glucose and fructose, but also methylglyoxal and other reactive carbonyl intermediates) react non-enzymatically with the free amino groups of lysine, arginine, and other amino acid residues on proteins. The reaction proceeds in stages: initial Schiff base formation, rearrangement to a stable Amadori product, and then further reactions over weeks to months to form the irreversible cross-linked end-products that we call AGEs.
AGEs accumulate progressively in long-lived proteins — collagen in skin and tendons, crystallins in the lens of the eye, vascular basement membranes, neuronal proteins. They cross-link extracellular matrix proteins, stiffening arteries (contributing to systolic hypertension and pulse-wave reflection), thickening basement membranes (contributing to diabetic nephropathy), causing lens opacification (contributing to senile cataracts), and triggering pro-inflammatory signaling through the RAGE (receptor for AGE) on macrophages and endothelial cells.
Carnosine inhibits AGE formation at multiple steps:
- Direct competition with protein lysine — the beta-alanine N-terminus of carnosine has a primary amino group that competes with protein lysine residues for the initial glycation reaction. Sugars react with carnosine rather than with protein, and the carnosine-sugar adduct is excreted.
- Methylglyoxal scavenging — methylglyoxal is a highly reactive intermediate in glycolysis that forms AGEs much faster than glucose itself. Carnosine reacts with methylglyoxal at high rate constants, intercepting it before it can damage proteins.
- Imidazole-aldehyde Michael addition — similar to the 4-HNE mechanism, the imidazole ring of carnosine forms stable adducts with reactive aldehyde intermediates in the AGE pathway.
- Carbonyl scavenging — carnosine is the most efficient endogenous "carbonyl scavenger" identified, with broad activity against the reactive carbonyl species that drive both glycation and lipid peroxidation.
The net effect is that carnosine acts as a sacrificial substrate for glycation, taking the chemical damage that would otherwise accumulate on long-lived structural proteins. This is the mechanistic basis for the strong epidemiological signal linking low plasma carnosine to accelerated tissue aging, and for the protective effect of CNDP1 genetic variants that raise plasma carnosine in diabetic patients.
Carnosine in Diabetic Complications
Type 2 diabetes is essentially a disease of accelerated glycation. Chronic hyperglycemia drives a several-fold increase in the rate of AGE formation throughout the body, producing the long-term complications that account for most diabetes-associated morbidity: diabetic nephropathy, diabetic retinopathy, diabetic neuropathy, accelerated atherosclerosis, and impaired wound healing.
Carnosine's anti-glycation activity makes it a logical therapeutic candidate, and the evidence base has been growing for two decades:
- Diabetic nephropathy — the most-studied indication. The CNDP1 (carnosinase) genetic polymorphism epidemiology was the original signal: patients homozygous for the "Mannheim allele" (5L/5L) of CNDP1 have low-activity serum carnosinase, elevated plasma carnosine, and approximately 50% reduced risk of nephropathy progression in type 2 diabetes. Multiple subsequent studies in European and Asian populations have replicated this finding.
- Diabetic retinopathy — carnosine reduces the formation of advanced glycation end-products in lens crystallins and retinal extracellular matrix. N-acetylcarnosine eye drops (the "carnosine cataract" product) have been studied in cataract progression with mixed results — positive signals in some Russian and Asian trials, less consistent results in Western trials.
- Glycemic control — small RCTs of oral carnosine (1-2 g/day) or beta-alanine (3-6 g/day) in pre-diabetic and type 2 diabetic patients have shown modest improvements in HbA1c (typically 0.2-0.5 percentage points), insulin sensitivity, and inflammatory markers. Effects are smaller than diet, exercise, or metformin, but the safety profile is excellent.
- Diabetic vascular disease — carnosine protects endothelial function in animal models and in some small human trials. The mechanism involves both AGE-reduction and direct preservation of nitric oxide signaling.
The current opinion is that carnosine and beta-alanine are reasonable adjuncts to standard diabetes management, particularly for patients with established complications or with the high-activity CNDP1 genotype that puts them at elevated nephropathy risk. They do not replace insulin, metformin, GLP-1 agonists, SGLT2 inhibitors, or other primary diabetes therapies, but they may meaningfully slow the rate of complication progression.
Age-Related Decline of Muscle Carnosine
Muscle carnosine concentration declines progressively with age in adults, with most studies showing a roughly 60% reduction between ages 25 and 70. The decline is parallel to (but distinct from) the better-known age-related declines in muscle mass, mitochondrial function, and creatine concentration.
Several factors contribute:
- Reduced beta-alanine supply — reduced meat intake with age, reduced gastric acid for protein digestion (atrophic gastritis is common after age 60), and reduced enterocyte transport capacity all reduce beta-alanine absorption.
- Reduced fast-twitch fiber mass — sarcopenia preferentially affects type II (high-carnosine) fibers, so total muscle carnosine declines partly as a function of overall fiber-type composition shift.
- Reduced carnosine synthase activity — aged muscle has reduced expression of carnosine synthase, possibly secondary to anabolic signaling decline.
- Increased oxidative carnosine consumption — aged tissue has higher baseline oxidative stress, which is "buffered" by carnosine via the mechanisms above, and this turnover depletes the pool faster than synthesis can replace it.
The functional consequence is reduced muscle buffering capacity, reduced peak power output, slower exercise recovery, and greater susceptibility to oxidative protein damage. Beta-alanine supplementation has been shown to reverse the carnosine decline in older adults to a substantial degree — typical regimens of 3-6 g/day for 12 weeks restore muscle carnosine to levels seen in much younger adults, with measurable improvements in functional capacity, exercise performance, and quality-of-life metrics in some trials.
For more on age-related sarcopenia, see our page on Sarcopenia, or our general Amino Acids landing page.
Anserine in Poultry and Fish, Balenine in Whale Muscle
Carnosine has two close relatives in the histidine-dipeptide family:
Anserine (beta-alanyl-N-pi-methylhistidine) is identical to carnosine except for a methyl group on the imidazole ring of the histidine moiety. It is the dominant histidine-dipeptide in poultry muscle (chicken breast, turkey breast), in salmonid fish (salmon, trout), and in tuna. Anserine has antioxidant and AGE-quenching activities similar to carnosine, but with somewhat different metal-chelation properties (the methyl group changes the imidazole pKa and electron density). Anserine is rapidly metabolized in human tissue to carnosine's constituents (beta-alanine and the methylated histidine), and the methylated histidine cannot be reincorporated into protein but is excreted in urine.
Anserine is the basis for the high carnosine-equivalent content of chicken breast, which delivers approximately 750 mg of combined carnosine + anserine per 100 g serving — far more than beef. This is one reason why poultry is consistently associated with skeletal-muscle benefits in older adult nutrition studies, beyond the simple high-quality-protein effect.
Balenine (beta-alanyl-N-tau-methylhistidine) is found in cetacean muscle (whales, dolphins) and certain reptile muscle. It is essentially absent from human dietary sources (whale meat consumption is now negligible in developed countries) but represents an interesting evolutionary variant — the muscle of large diving mammals contains extraordinary concentrations of these histidine dipeptides, contributing to dive-tolerance through enhanced pH buffering during the prolonged anaerobic exercise of breath-holding dives.
The pattern across species is consistent: muscle that needs to operate at high intensity under acidotic conditions (sprint muscle of mammals, fish on rapid escape responses, diving muscle of whales) is rich in histidine dipeptides. This has been one of the more elegant evolutionary signals for the importance of these compounds in real-world athletic performance.
Brain Carnosine and Polaprezinc (Zinc-Carnosine)
Beyond muscle, carnosine has clinically relevant activities in two other compartments:
Brain (particularly olfactory bulb). Olfactory bulb has the highest carnosine concentration of any brain region (1-3 mmol/kg wet), where it acts as a co-released signaling molecule from olfactory sensory neurons and provides antioxidant protection in the high-turnover olfactory epithelium. Cerebellar Purkinje cells and certain hypothalamic nuclei also express moderate carnosine. The role in cognition and neurodegeneration is an active research area — carnosine and its zinc complex have been studied as adjuncts in autism spectrum disorder (some signals in small RCTs), in Alzheimer's disease (mechanistic plausibility for amyloid-beta interaction), and in Parkinson's disease (animal-model evidence for protection of dopaminergic neurons).
Polaprezinc (zinc-L-carnosine). A chelate compound of zinc and carnosine, marketed in Japan since 1994 (brand name Promac) as a treatment for gastric ulcer disease, and now widely used internationally as an over-the-counter supplement for gut health. The chelate has dual mechanisms: the carnosine moiety provides direct mucosal antioxidant protection and quenches the lipid peroxidation that contributes to gastric mucosal damage, while the zinc provides additional epithelial barrier support and antimicrobial activity against Helicobacter pylori (the dominant cause of peptic ulcer disease).
RCT evidence supports polaprezinc 75 mg twice daily as adjunct therapy in H. pylori eradication regimens (modestly improved eradication rates), in NSAID-induced gastritis (significantly reduced gastric mucosal damage), and in chemotherapy-induced oral mucositis (modest reduction in pain and ulcer severity). The compound has an excellent safety profile and is one of the more useful applications of carnosine pharmacology in routine clinical practice.
Supplementation: Histidine, Beta-Alanine, or Carnosine?
The practical question for anyone wanting to raise muscle carnosine concentration is: which precursor to supplement?
Direct carnosine supplementation is largely ineffective for raising muscle carnosine in healthy adults. Oral carnosine is rapidly hydrolyzed by intestinal and serum carnosinase to its constituent beta-alanine and histidine within minutes. The intact carnosine that reaches the systemic circulation is negligible. There is some evidence for direct carnosine effects on gut epithelium and on certain mucosal sites (polaprezinc above is a special case), but for muscle and systemic effects, supplementing carnosine itself is a poor strategy.
Histidine supplementation does not raise muscle carnosine in healthy adults, because histidine is essentially never rate-limiting in carnosine synthesis (see the kinetic discussion above). The exception is in chronic kidney disease, where dialysis depletes plasma histidine; in that setting, histidine supplementation does support both hemoglobin synthesis and muscle carnosine, but the indication is the underlying histidine deficiency, not the carnosine elevation per se.
Beta-alanine supplementation is the established protocol for raising muscle carnosine. Typical regimen: 3-6 g/day for 8-12 weeks. The dose is split into multiple smaller doses (e.g., 800 mg four times daily) to minimize the "tingling" sensation (paresthesia from MRGPRD activation) that is unpleasant but harmless. Sustained-release beta-alanine formulations reduce the paresthesia effectively.
Expected results from a 12-week beta-alanine loading:
- Muscle carnosine increases 40-80% from baseline
- High-intensity exercise capacity (1-4 minute duration) improves 2-3% on average
- Buffering capacity in working muscle increases meaningfully
- Lean body mass changes are variable and typically modest
- Reduced markers of exercise-induced oxidative stress in some studies
Beta-alanine is one of the few sports-nutrition supplements with sufficient RCT evidence to be on the International Olympic Committee's "Strong" evidence list for ergogenic effect (alongside creatine, caffeine, sodium bicarbonate, and nitrate).
Combined beta-alanine + histidine is sometimes promoted as superior to beta-alanine alone, but the kinetic argument suggests no additional benefit unless the individual is in a histidine-deficient state.
Key Research Papers
- Gulewitsch W, Amiradzibi S (1900). Ueber das Carnosin, eine neue organische Base des Fleischextractes. Berichte der deutschen chemischen Gesellschaft. The original isolation paper. — PubMed
- Boldyrev AA et al. (2013). Physiology and pathophysiology of carnosine. Physiological Reviews. The definitive modern review. — PubMed
- Harris RC et al. (2006). The absorption of orally supplied beta-alanine and its effect on muscle carnosine synthesis in human vastus lateralis. Amino Acids. — PubMed
- Hill CA et al. (2007). Influence of beta-alanine supplementation on skeletal muscle carnosine concentrations and high intensity cycling capacity. Amino Acids. — PubMed
- Hipkiss AR (2009). Carnosine and its possible roles in nutrition and health. Advances in Food and Nutrition Research. — PubMed
- Aldini G et al. (2002). Identification of histidyl-dipeptides as endogenous reactive carbonyl species scavengers. Biochemical and Biophysical Research Communications. — PubMed
- Janssen B et al. (2005). Carnosine as a protective factor in diabetic nephropathy: association with a leucine repeat of the carnosinase gene CNDP1. Diabetes. The Mannheim allele paper. — PubMed
- Sale C et al. (2010). Effect of beta-alanine supplementation on muscle carnosine concentrations and exercise performance. Amino Acids. — PubMed
- Stegen S et al. (2014). Plasma carnosine, but not muscle carnosine, attenuates high-fat-diet-induced metabolic stress. Applied Physiology, Nutrition, and Metabolism. — PubMed
- Mayuko M et al. (2008). Effect of polaprezinc (zinc-L-carnosine complex) on Helicobacter pylori eradication. Internal Medicine. — PubMed
- Trexler ET et al. (2015). International society of sports nutrition position stand: beta-alanine. Journal of the International Society of Sports Nutrition. — PubMed
- Hipkiss AR (2020). Possible benefit of dietary carnosine towards depressive disorders. Aging and Disease. — PubMed
PubMed Topic Searches
- PubMed: Carnosine antioxidant activity
- PubMed: Beta-alanine and exercise performance
- PubMed: Carnosine and AGE quenching
- PubMed: Anserine in poultry and fish
- PubMed: Polaprezinc for gastric disease
Connections
- Histidine Overview
- Histidine Benefits Hub
- Histidine, Histamine, and Allergy
- Histidine for Hemoglobin
- Histidine for Wound and Joint Health
- All Amino Acids
- Beta-Alanine
- Zinc
- Copper
- Oxidative Stress
- Type 2 Diabetes
- Diabetic Nephropathy
- Cataracts
- Chicken (Anserine Source)
- Tuna (Anserine Source)