Glycine for Collagen Synthesis
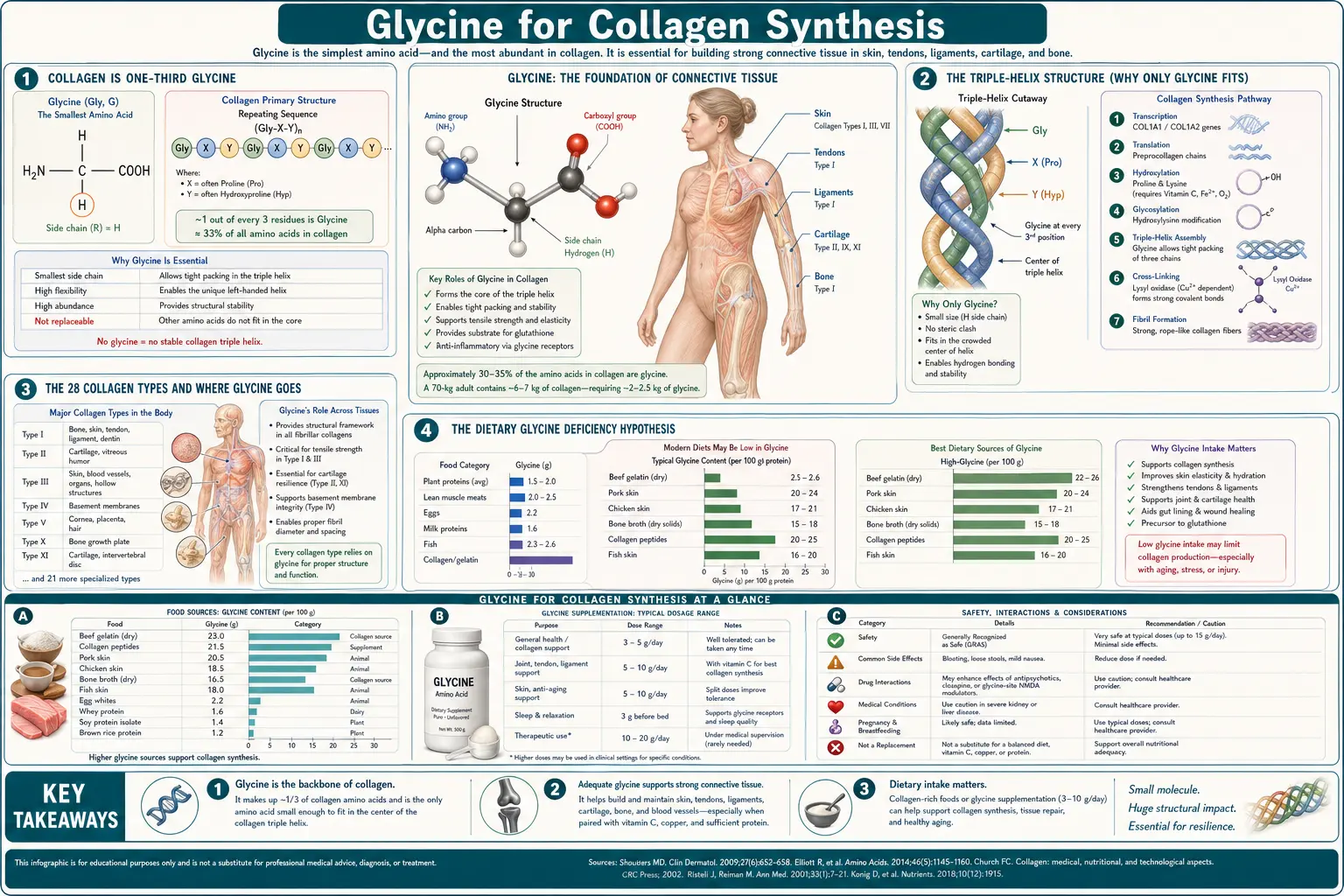
Collagen is the most abundant protein in the human body — roughly 30% of total body protein and the structural backbone of skin, bone, cartilage, tendons, ligaments, blood-vessel walls, fascia, and the gut lining. Every third amino acid in collagen's repeating triple-helix is glycine. No other amino acid is small enough to fit. The modern Western diet, which strips muscle meat from the connective-tissue cuts that traditional cuisines used (oxtail, marrow bones, skin, tendons, hooves, knuckle joints), supplies dramatically less glycine than ancestral and traditional patterns. This deep-dive walks through the structural chemistry of why glycine is mandatory at every third position, the dietary-glycine-deficiency hypothesis advanced by Meléndez-Hevia and McCarty, the wound-healing literature showing accelerated recovery with glycine supplementation, the GlyNAC aging-reversal work of Rajagopal Sekhar at Baylor, and the practical hierarchy of collagen-rich foods.
Table of Contents
- Collagen Is One-Third Glycine
- The Triple-Helix Structure (Why Only Glycine Fits)
- The 28 Collagen Types and Where Glycine Goes
- The Dietary Glycine Deficiency Hypothesis
- Muscle Meat vs Whole-Animal Eating
- Wound Healing and Surgical Recovery
- Skin Aging and Collagen Peptide Trials
- Joints, Cartilage, and Tendon Repair
- Bone Broth, Gelatin, and Collagen Peptides
- GlyNAC, Sekhar, and the Aging Connection
- Cofactors: Vitamin C, Copper, and Lysine
- Practical Dosing
- Key Research Papers
- Connections
- Featured Videos
Collagen Is One-Third Glycine
Collagen is unique among human proteins in its amino-acid composition. A typical protein contains all twenty standard amino acids in roughly random proportion. Collagen does not. Its primary sequence is dominated by a repeating Gly-X-Y tripeptide motif, where X is most often proline and Y is most often hydroxyproline (a post-translationally modified form of proline). This repeat occupies roughly 95% of the central triple-helical domain of every collagen polypeptide.
The arithmetic consequence: glycine accounts for approximately 33% of collagen's amino acid residues by count — one out of every three. Proline and hydroxyproline together account for roughly another 22%. The remaining 45% of positions are filled by the other 17 amino acids in roughly conventional proportions.
Because collagen is approximately 30% of total body protein, and glycine is 33% of collagen, glycine in collagen alone accounts for roughly 10% of total body protein-bound amino acids. No other amino acid is so heavily concentrated in a single protein class, and no other amino acid sees turnover demand on the scale of daily collagen remodeling.
The total daily glycine demand for adult collagen turnover alone has been estimated at 10–12 grams per day, against an endogenous synthesis capacity of only about 3 g/day and typical Western dietary glycine intake of 3–5 g/day. The gap is the basis for the dietary-glycine-deficiency hypothesis discussed below.
The Triple-Helix Structure (Why Only Glycine Fits)
Collagen's defining structural feature is the triple helix — three left-handed polyproline-II-type polypeptide chains wound around each other in a right-handed superhelix. This structure was first proposed by Pauling and Corey and refined by Ramachandran in the 1950s. The triple helix gives collagen its characteristic tensile strength — a single collagen fibril can withstand more force per cross-sectional area than steel.
The geometry of the triple helix imposes an absolute structural requirement: the residue at the innermost position of each turn must have a hydrogen atom as its side chain. That position is the third residue of every Gly-X-Y triplet. Any side chain larger than a single hydrogen physically cannot fit into the cramped axial space of the triple helix. Only glycine, with its single-H side chain, qualifies.
This is not a matter of preference or efficiency — it is a hard structural constraint. The mutations that produce osteogenesis imperfecta (brittle bone disease) almost universally involve substitution of a larger amino acid for one of the obligate glycines in collagen type I. Even a single glycine-to-cysteine substitution in a 1,000-residue collagen chain can produce a clinically dramatic phenotype, because the substitution kinks the triple helix at that point and disrupts the assembly of higher-order collagen fibrils. Similar glycine-substitution mutations in collagen type III produce vascular Ehlers-Danlos syndrome, in type IV produce Alport syndrome (kidney), and in type II produce chondrodysplasia phenotypes.
The genetic disorders prove the structural necessity. Glycine is not a "preferred" residue at the third position of the triplet — it is the only residue that allows the triple helix to form at all.
The 28 Collagen Types and Where Glycine Goes
There are at least 28 distinct collagen types in the human body, encoded by 43 collagen genes. They are grouped into fibril-forming collagens (types I, II, III, V, XI), network-forming collagens (type IV in basement membranes, type X in cartilage), fibril-associated collagens (FACIT family), and others. Each is built from the same Gly-X-Y repeat pattern, so each requires the same one-in-three glycine residues.
The dominant types by mass are:
- Type I collagen (~90% of total body collagen) — the structural collagen of skin (dermis), bone, tendon, ligament, fascia, and the cornea. Two alpha-1(I) chains plus one alpha-2(I) chain per triple helix.
- Type II collagen — the principal collagen of hyaline cartilage in joints, the nucleus pulposus of intervertebral discs, and the vitreous humor of the eye.
- Type III collagen — co-distributed with type I in skin, blood vessel walls, and the reticular framework of liver, spleen, and lymph node. Particularly important in wound healing, where it forms the early granulation tissue before type I deposition takes over.
- Type IV collagen — the network collagen of basement membranes underlying every epithelial and endothelial layer. The kidney glomerular basement membrane is largely type IV.
- Type V collagen — minor collagen co-assembled with type I in skin, bone, and cornea; regulates fibril diameter.
Every one of these uses the same Gly-X-Y motif and draws from the same hepatic and tissue glycine pool. When dietary glycine is limited, the tissues with the highest collagen turnover — skin, gut lining, bone-remodeling sites, healing wounds — show the deficit first.
The Dietary Glycine Deficiency Hypothesis
The proposition that modern humans are chronically glycine-deficient was advanced most explicitly by Enrique Meléndez-Hevia and colleagues at the University of La Laguna in a 2009 paper in Journal of Biosciences titled "A weak link in metabolism: the metabolic capacity for glycine biosynthesis does not satisfy the need for collagen synthesis." The argument:
- Total glycine demand for collagen turnover plus all other glycine-using pathways (glutathione, creatine, heme, bile acids, nucleotides, phase II detoxification) is approximately 15 g/day in a healthy adult.
- Endogenous glycine synthesis through serine hydroxymethyltransferase and threonine catabolism produces approximately 3 g/day.
- Typical modern Western dietary glycine intake is 1.5–3 g/day (muscle meat is glycine-poor relative to connective tissue).
- The shortfall is approximately 9–10 g/day, met by reducing collagen turnover, reducing glutathione synthesis, reducing detoxification capacity, or some combination.
The same argument was independently made by Mark McCarty in a series of papers published from 2014 onward, arguing that the modern glycine shortfall contributes to accelerated aging, vascular disease, sarcopenia, slow wound healing, and the metabolic syndrome cluster. McCarty's framing emphasizes that the daily deficit is sub-clinical — it does not produce frank deficiency in the way that scurvy proves a vitamin C deficit — but compounds over decades into measurable tissue consequences.
The deficiency hypothesis is not universally accepted. Conventional nutrition recognizes glycine as a "non-essential" amino acid because the body can synthesize it from serine, and serine can in turn be synthesized from glucose plus a nitrogen source. By that strict definition, glycine cannot be deficient because the body can always make more. The Meléndez-Hevia and McCarty counterargument is that the maximum biosynthetic capacity falls short of the maximum metabolic demand, which is a different question than whether the amino acid is technically essential.
The clinical evidence supporting the hypothesis is mostly indirect: epidemiologic associations of low plasma glycine with metabolic syndrome, type 2 diabetes, sarcopenia, and cardiovascular disease; observational data on accelerated skin aging in low-glycine populations; and intervention studies (GlyNAC, collagen peptides) showing measurable benefit when glycine is supplemented above typical dietary intake. Whether you call this "deficiency" or "insufficiency" or "suboptimal intake," the practical recommendation is the same: increase intake.
Muscle Meat vs Whole-Animal Eating
The single largest dietary determinant of glycine intake is whether one eats whole-animal (including connective tissues, skin, organ meats, and bone-derived broths) or muscle-only (boneless skinless chicken breast, lean ground beef, fish fillets). The amino-acid composition difference is substantial:
- Pork or beef shoulder, slow-cooked with connective tissue: ~3–5 g glycine per 100 g
- Chicken with skin: ~2–3 g glycine per 100 g
- Beef tendon, oxtail, knuckle: ~4–6 g glycine per 100 g
- Boneless skinless chicken breast: ~1.2 g glycine per 100 g
- Lean ground beef (90/10): ~1.5 g glycine per 100 g
- Skinless salmon fillet: ~1.4 g glycine per 100 g
- Bone broth, well-prepared: ~2–5 g glycine per 250 mL serving
- Hydrolyzed collagen peptides: ~3.3 g glycine per 10 g serving
Modern industrial agriculture has further widened the gap. Mass-produced meat is increasingly delivered as boneless, skinless, trimmed cuts — convenient, quickly cooked, and entirely shorn of the collagen-rich tissues that traditional kitchens prized. Stocks and broths take 12–24 hours to prepare properly and have largely been replaced by bouillon cubes that contain no real gelatin. The shift parallels (and probably contributes to) the apparent rise in glycine-deficiency-sensitive conditions.
The Meléndez-Hevia argument concludes that recovering ancestral whole-animal eating — or, if that is impractical, supplementing with collagen peptides or free glycine — is the simplest dietary correction available.
Wound Healing and Surgical Recovery
Wound healing places dramatically elevated demand on collagen synthesis. Type III collagen lays down the initial granulation tissue within 3–5 days of wounding. Type I collagen then progressively replaces type III over weeks to months as the wound matures into a stable scar. The total mass of collagen synthesized at a major surgical or traumatic wound can equal weeks of normal whole-body collagen turnover.
Glycine supplementation has been studied in several wound-healing contexts:
- Diabetic foot ulcers. Several small trials of glycine plus arginine plus antioxidants have reported faster healing and reduced inflammation compared to standard care alone.
- Pressure ulcers. Collagen-peptide supplementation in elderly nursing-home residents with stage 2–4 pressure injuries has shown improved healing rates in randomized trials.
- Burns. Glycine plus arginine plus glutamine (a common immunonutrition combination) has been studied in burn-unit protocols; evidence of benefit is suggestive but not conclusive.
- Postoperative recovery. Small studies suggest that pre-operative collagen-peptide supplementation may reduce postoperative wound complications, particularly in patients undergoing abdominal surgery, but the trials are small and underpowered for hard outcomes.
- Sports-related soft-tissue injury. The Shaw et al. 2017 trial showed that 15 g of gelatin (with vitamin C) consumed an hour before exercise increased collagen synthesis markers measured in serum, suggesting a window for targeted nutritional support of injury repair.
The mechanistic case is straightforward: collagen synthesis at a wound is rate-limited by glycine, proline, and hydroxyproline availability (and, indirectly, by vitamin C, which is the cofactor for prolyl-hydroxylase). Providing more of those substrates increases the local capacity for collagen deposition. Whether this translates to clinically meaningful faster healing depends on baseline status, wound type, and concurrent conditions.
Skin Aging and Collagen Peptide Trials
Skin collagen content declines progressively with age. Type I collagen in the dermis decreases by approximately 1% per year after age 20 in women and slightly less in men. Postmenopausal estrogen decline accelerates the loss — women lose roughly 30% of dermal collagen in the first 5 years after menopause. The clinical consequences are visible: thinner skin, more wrinkles, slower wound healing, more bruising, and reduced skin elasticity.
The randomized-controlled-trial evidence for oral collagen peptides as an intervention for skin aging is now substantial. The most-cited trials include:
- Proksch et al. 2014 (Skin Pharmacology and Physiology). Bovine collagen peptides (Verisol, 2.5 g daily for 8 weeks) improved skin elasticity in postmenopausal women compared to placebo.
- Asserin et al. 2015 (Journal of Cosmetic Dermatology). 10 g daily of bioactive collagen peptides for 8 weeks improved skin hydration and dermal collagen density measured by ultrasound.
- Kim et al. 2018 (Nutrients). 1 g daily of fish-derived collagen peptides for 12 weeks improved skin moisture, elasticity, and wrinkle parameters in middle-aged women.
- de Miranda et al. 2021 meta-analysis (International Journal of Dermatology). Pooled analysis of 19 trials confirmed significant improvements in skin elasticity, hydration, and wrinkle reduction with oral collagen peptide supplementation across 2.5–10 g daily doses.
The mechanism is partly nutritional substrate provision (glycine, proline, hydroxyproline-rich peptides as raw material for new collagen synthesis) and partly signaling — some collagen-derived dipeptides (Pro-Hyp, Hyp-Gly) appear to act as low-grade signaling molecules that stimulate fibroblast proliferation and matrix deposition. Free-form glycine alone has been studied less extensively for skin outcomes specifically, but the substrate logic suggests it should provide a fraction of the benefit at lower cost.
Joints, Cartilage, and Tendon Repair
Articular cartilage is type II collagen embedded in an aggrecan-rich proteoglycan matrix. Cartilage has no blood supply and depends on diffusion through the joint synovial fluid for both nutrient delivery and waste removal — which is why cartilage damage heals so slowly and why osteoarthritis progression is so difficult to reverse.
Tendons and ligaments are dense bundles of type I collagen fibrils aligned with the direction of tensile load. They turn over more slowly than skin but face equivalent mechanical demands.
Several lines of evidence support glycine and collagen-peptide supplementation for joint and tendon health:
- Clark et al. 2008 (Current Medical Research and Opinion). 10 g daily of collagen hydrolysate for 24 weeks in athletes with activity-related joint pain reduced pain scores significantly compared to placebo.
- Bruyere et al. 2012 (Complementary Therapies in Medicine). 12 g daily of collagen hydrolysate for 6 months in patients with osteoarthritis reduced joint discomfort.
- Shaw et al. 2017 (American Journal of Clinical Nutrition). 15 g of gelatin plus 50 mg vitamin C consumed an hour before 6 minutes of jump-rope skipping doubled serum markers of collagen synthesis, suggesting timed intake at peak load is uniquely beneficial for tendon and ligament adaptation.
- Praet et al. 2019 (Nutrients). Specific bioactive collagen peptides studied for Achilles tendinopathy showed improvements in pain and function.
The Shaw findings have launched a small movement among athletes and physical therapists to time gelatin/collagen intake to peak training stress, on the rationale that the 30–60 minute post-ingestion window is when fibroblasts in tendon and cartilage are receiving the highest concentration of collagen substrate and (presumably) depositing the most new collagen.
Bone Broth, Gelatin, and Collagen Peptides
The three practical dietary vehicles for collagen and glycine are bone broth (traditional, food), gelatin (intact collagen powder, food-grade), and hydrolyzed collagen peptides (enzymatically pre-digested, dietary-supplement-style).
Bone broth is made by simmering animal bones, joints, and connective-tissue scraps in water for 8–24 hours, often with a splash of acid (vinegar or wine) to facilitate mineral extraction. The long simmer hydrolyzes the bone collagen into soluble gelatin, which makes the cooled broth gel-like. A good homemade beef-bone broth yields 2–5 grams of glycine per 250 mL cup, plus minerals, glycosaminoglycans, and other connective-tissue components. Commercial bone broths vary widely — many are essentially flavored stock with minimal gelatin; quality-marked premium brands gel when refrigerated, which is the simplest test of meaningful collagen content.
Gelatin is partially hydrolyzed collagen, sold as a powder or as sheets, that gels when dissolved in hot water and cooled. It is the active ingredient in homemade jellies, panna cotta, marshmallows, gummies, and aspic. Gelatin does not dissolve in cold liquids — it must be "bloomed" by sprinkling on cold water and then warmed. About 33% glycine by weight, so 10 g of gelatin delivers ~3.3 g of glycine.
Hydrolyzed collagen peptides are gelatin further broken down by enzymatic digestion into shorter peptides (typically 3–5 kDa). They dissolve in both cold and hot liquids, do not gel, and absorb more rapidly. Same 33% glycine content as gelatin. Most commercial collagen-peptide products are bovine-hide- or fish-skin-derived. A 10–20 g daily serving delivers 3–7 g of glycine plus the full collagen amino-acid profile.
Free-form glycine powder is the cheapest and purest option, delivering 100% glycine by weight. It does not provide proline, hydroxyproline, or any of the other amino acids that collagen peptides contribute. For sleep, aspirin metabolism, and glutathione purposes, free glycine alone is sufficient. For collagen synthesis specifically, collagen peptides or gelatin probably have a small edge because they provide the proline and hydroxyproline substrates as well.
GlyNAC, Sekhar, and the Aging Connection
Rajagopal Sekhar at Baylor College of Medicine has led a research program for nearly two decades on the role of glycine, cysteine, and glutathione in aging. His central finding is that older adults have markedly lower red-blood-cell and tissue glutathione than younger adults, that this glutathione deficit is largely driven by inadequate substrate supply of both glycine and cysteine, and that supplementing both amino acids together (the "GlyNAC" combination of glycine plus N-acetylcysteine) substantially restores glutathione, reduces oxidative stress markers, improves mitochondrial function, and improves a wide range of clinical biomarkers in older adults.
The collagen-synthesis story is the parallel narrative on the same substrate base. Glycine going into glutathione is glycine not going into collagen, and vice versa. The aging tissues that show declining collagen integrity (skin thinning, cartilage loss, bone weakening, vascular wall stiffening) are also the tissues with declining glutathione and rising oxidative stress markers. The Sekhar GlyNAC trials — covered in detail on the Glutathione Synthesis page — document multi-system improvements that are best explained by simultaneous correction of both the antioxidant deficit and the structural-substrate deficit.
The practical implication for the collagen story is that addressing glycine adequacy in older adults supports both the structural collagen turnover that maintains tissue integrity and the antioxidant capacity that defends against the oxidative damage that drives further collagen cross-linking and stiffening.
Cofactors: Vitamin C, Copper, and Lysine
Glycine is necessary but not sufficient for collagen synthesis. Several cofactors are essential at the enzymatic steps that produce mature collagen fibrils:
- Vitamin C (ascorbate) — the cofactor for prolyl-4-hydroxylase and lysyl-hydroxylase, the enzymes that post-translationally hydroxylate proline and lysine residues in the nascent collagen polypeptide. Hydroxyproline and hydroxylysine stabilize the triple helix; without them, the collagen molecule unwinds and is degraded. This is the molecular basis of scurvy, in which dietary vitamin C deficiency leads to defective collagen, bleeding gums, easy bruising, poor wound healing, and the dramatic re-opening of old wounds that historically gave scurvy its diagnostic profile in sailors. Maintain at least 200 mg vitamin C daily for adequate collagen support; some integrative protocols use 500–1,000 mg.
- Copper — the cofactor for lysyl oxidase, the enzyme that initiates the cross-linking of mature collagen fibrils via deamination of lysine and hydroxylysine residues. Copper deficiency produces weak, poorly cross-linked collagen and contributes to vascular fragility and bruising. See the Copper page for context.
- Lysine — one of the most common residues at the Y position of the Gly-X-Y triplet, particularly in skin and tendon collagen. Adequate dietary lysine supports collagen synthesis.
- Proline — the second most abundant collagen amino acid (X position), supplied directly by collagen peptides or by endogenous synthesis from glutamate.
- Iron — the prolyl-hydroxylase and lysyl-hydroxylase enzymes contain non-heme iron at their active sites. Severe iron deficiency impairs collagen hydroxylation.
- Zinc — cofactor for matrix metalloproteinases that remodel collagen during turnover and wound healing; also supports many other steps of tissue repair.
- Manganese and silicon — supporting roles in connective-tissue glycosaminoglycan biosynthesis.
The practical implication: a glycine intervention for collagen support should normally be paired with adequate vitamin C, lysine (easily covered by ordinary protein intake), and a basic mineral baseline. A single-nutrient glycine supplement in someone with marginal vitamin C status will not produce the expected wound-healing or skin-aging benefit.
Practical Dosing
Collagen-synthesis-targeted glycine dosing differs from sleep-targeted dosing in both quantity and timing:
- Daily maintenance. 5–10 g of glycine per day (from food + supplements combined), split through the day or taken once. For most adults, one of the following meets the target:
- 10–20 g of collagen peptides daily (provides ~3.3–7 g glycine plus proline and hydroxyproline)
- 1 cup of well-made bone broth daily (provides ~2–5 g glycine plus broader connective-tissue nutrients)
- 5 g of free glycine powder daily plus normal dietary protein
- Active wound healing or post-surgical recovery. Increase to 15–20 g of collagen peptides daily plus 500–1,000 mg vitamin C, divided through the day, for the active healing phase (typically 2–6 weeks).
- Tendon and ligament training adaptation. Following the Shaw protocol, 15 g of gelatin or collagen peptides plus 50–100 mg vitamin C consumed 30–60 minutes before a tendon-loading session (jumping, sprinting, resistance training).
- Skin-aging intervention. 2.5–10 g of bioactive collagen peptides daily for at least 8–12 weeks before assessing effect. Sustained intake is required — benefits regress within months of stopping.
- GlyNAC for aging. Per the Sekhar protocol, 1.2–3.6 g of glycine plus equivalent N-acetylcysteine daily; see the Glutathione Synthesis page for details.
For most users, 10–20 g of collagen peptides daily is the simplest and most effective single intervention because it provides glycine, proline, and hydroxyproline together in roughly the proportions that collagen synthesis needs. Free glycine powder is cheaper but provides only the glycine fraction. Bone broth is the most traditional option but the most variable in actual collagen content.
Key Research Papers
- Meléndez-Hevia E, De Paz-Lugo P, Cornish-Bowden A, Cárdenas ML (2009). A weak link in metabolism: the metabolic capacity for glycine biosynthesis does not satisfy the need for collagen synthesis. Journal of Biosciences. — PubMed
- McCarty MF, O'Keefe JH, DiNicolantonio JJ (2018). Dietary glycine is rate-limiting for glutathione synthesis and may have broad potential for health protection. Ochsner Journal. — PubMed
- Shaw G, Lee-Barthel A, Ross ML, Wang B, Baar K (2017). Vitamin C-enriched gelatin supplementation before intermittent activity augments collagen synthesis. American Journal of Clinical Nutrition. — PubMed
- Proksch E, Segger D, Degwert J, et al. (2014). Oral supplementation of specific collagen peptides has beneficial effects on human skin physiology. Skin Pharmacology and Physiology. — PubMed
- Asserin J, Lati E, Shioya T, Prawitt J (2015). The effect of oral collagen peptide supplementation on skin moisture and the dermal collagen network. Journal of Cosmetic Dermatology. — PubMed
- Clark KL, Sebastianelli W, Flechsenhar KR, et al. (2008). 24-Week study on the use of collagen hydrolysate as a dietary supplement in athletes with activity-related joint pain. Current Medical Research and Opinion. — PubMed
- Kumar S, Sugihara F, Suzuki K, et al. (2015). A double-blind, placebo-controlled, randomised, clinical study on the effectiveness of collagen peptide on osteoarthritis. Journal of the Science of Food and Agriculture. — PubMed
- Kang MC, Yumnam S, Kim SY (2018). Oral intake of collagen peptide attenuates ultraviolet B irradiation-induced skin dehydration in vivo. Marine Drugs. — PubMed
- Persikov AV, Ramshaw JA, Brodsky B (2005). Prediction of collagen stability from amino acid sequence. Journal of Biological Chemistry. — PubMed
- Sekhar RV, McKay SV, Patel SG, et al. (2011). Glutathione synthesis is diminished in patients with uncontrolled diabetes and restored by dietary supplementation with cysteine and glycine. Diabetes Care. — PubMed
- Marini JC, Forlino A, Bachinger HP, et al. (2017). Osteogenesis imperfecta. Nature Reviews Disease Primers. — PubMed
- de Miranda RB, Weimer P, Rossi RC (2021). Effects of hydrolyzed collagen supplementation on skin aging: a systematic review and meta-analysis. International Journal of Dermatology. — PubMed
- Sekhar RV (2021). GlyNAC supplementation improves glutathione deficiency, oxidative stress, mitochondrial dysfunction, inflammation, insulin resistance, endothelial dysfunction, genotoxicity, muscle strength, and cognition in older adults: a randomized clinical trial. Clinical and Translational Medicine. — PubMed
PubMed Topic Searches
- PubMed: glycine and collagen synthesis
- PubMed: collagen peptides and skin aging
- PubMed: gelatin and tendon adaptation
- PubMed: bone broth collagen content
- PubMed: collagen and wound healing
Connections
- Glycine Overview
- Glycine Benefits Hub
- Glycine for Sleep and Relaxation
- Glycine for Glutathione Synthesis
- Glycine and Aspirin Metabolism
- Proline
- Lysine
- Vitamin C
- Copper
- Zinc
- Manganese
- Bone Broth
- Collagen
- Organ Meats
- Osteoarthritis
- Osteoporosis
- Skin Aging
- Wound Healing
- All Amino Acids