Aspartic Acid — Benefits Deep Dive
Aspartic acid is one of the most strategically placed metabolites in human biochemistry. The same simple four-carbon dicarboxylic amino acid serves as an excitatory neurotransmitter at NMDA receptors, the only nitrogen donor for the argininosuccinate synthetase step of the urea cycle, an anaplerotic refill for the TCA cycle, a gluconeogenic substrate, and the N1 atom of every purine nucleotide built de novo. Its chiral D-isomer takes a completely different path — concentrating in the pituitary, testes, and ovaries, where it modulates testosterone and the broader endocrine cascade. Four deep-dive pages below unpack the major biological roles, the supporting clinical evidence, and the practical implications for patients considering aspartate-containing supplements or facing diseases that disrupt aspartate metabolism.
Deep-Dive Articles
Neurotransmission & the NMDA Receptor
L-aspartate as the second major excitatory neurotransmitter (alongside glutamate), how the NMDA receptor functions as a coincidence-detection switch for long-term potentiation and memory formation, the distinction between L- and D-aspartate, the excitotoxicity cascade that drives neuronal death in stroke and traumatic brain injury, and the clinical role of NMDA antagonists like ketamine and memantine.
The Urea Cycle
Aspartate as the obligatory nitrogen donor at the argininosuccinate synthetase (ASS1) step of the urea cycle, the five urea-cycle enzyme defects (CPS1, OTC, ASS1, ASL, ARG1) plus the cofactor and transporter disorders (NAGS, HHH, citrin/CTLN2), the malate-aspartate shuttle bridging cytosol and mitochondria, and the management of acute and chronic hyperammonemia including LOLA in hepatic encephalopathy.
D-Aspartate & Testosterone
D-aspartate (the chiral isomer) and its accumulation in the anterior pituitary, testes, and ovaries; modulation of LH, FSH, growth hormone, and testosterone release; Topo and D'Aniello's controversial 2009 Italian RCT showing a 42% testosterone increase; Willoughby and Leutholtz's pivotal 2013 failed replication; the multi-million-dollar supplement market that persists despite mixed evidence; and what the data actually support.
Energy Production
Aspartate at the strategic crossroads of cellular energy: TCA-cycle anaplerosis via the aspartate-oxaloacetate equilibrium, the malate-aspartate shuttle moving cytosolic NADH into mitochondria for ATP synthesis, the gluconeogenic pathway during fasting, the purine nucleotide cycle in exercising muscle, and the historical (and modern) story of potassium-magnesium aspartate as an ergogenic supplement.
Table of Contents
- Deep-Dive Articles
- Why Aspartic Acid Produces Effects Across Many Systems
- Research Papers: Neurotransmission & NMDA
- Research Papers: Urea Cycle & Hyperammonemia
- Research Papers: D-Aspartate & Endocrine Function
- Research Papers: Energy Production & Mitochondrial Function
- Research Papers: Cross-Cutting (Mechanism, Status, Safety)
- External Authoritative Resources
- Connections
- Featured Videos
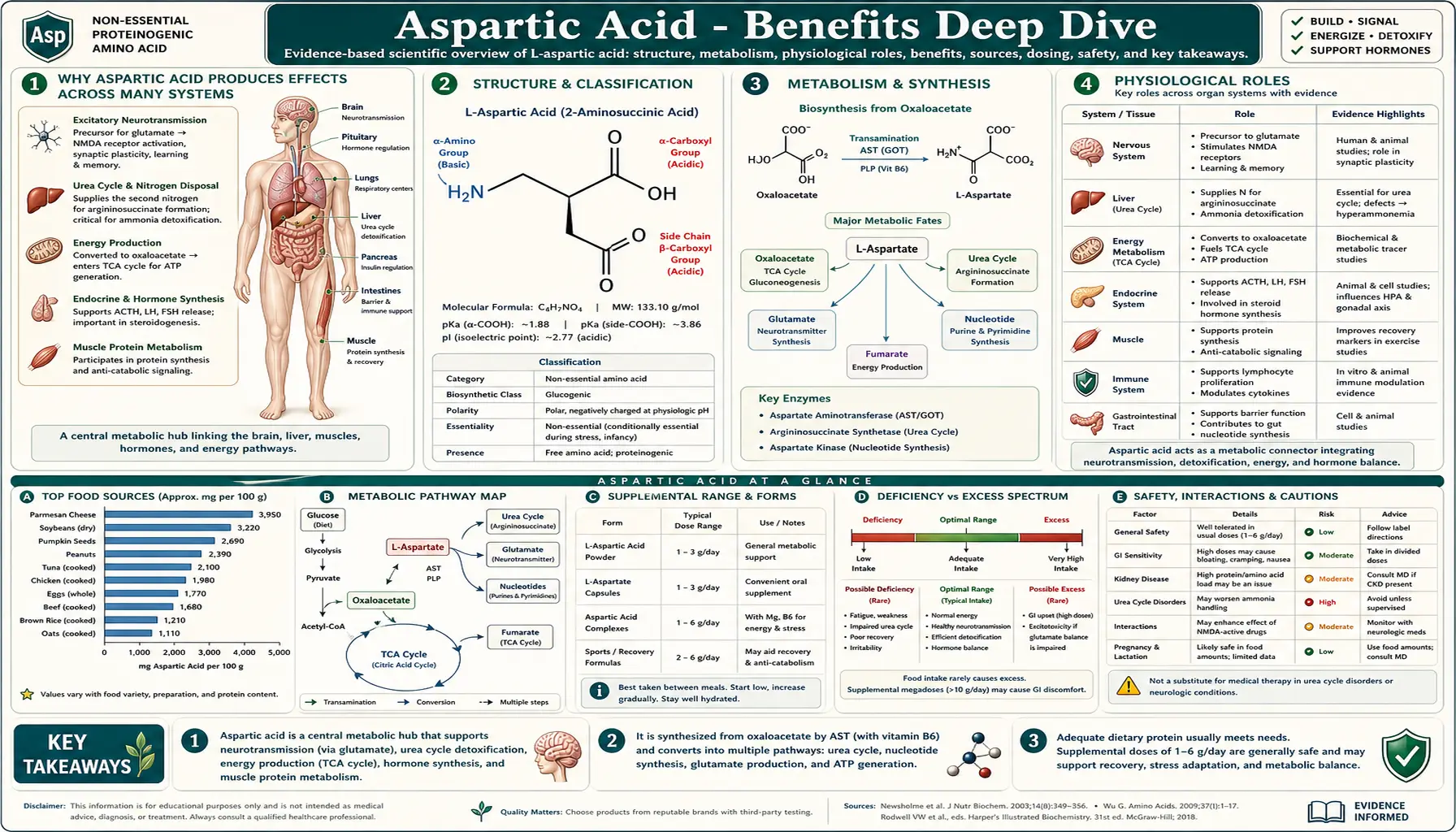
Why Aspartic Acid Produces Effects Across Many Systems
Most amino acids act through one principal mechanism — serving as protein building blocks plus, in some cases, one or two specialized metabolic roles. Aspartic acid is unusual because it operates through four fundamentally different mechanisms in two different molecular forms, and each maps to a distinct category of clinical effect.
- Excitatory neurotransmission via NMDA receptor activation (L-aspartate) — L-aspartate is released at glutamatergic synapses throughout the central nervous system, binds the NMDA receptor as a co-agonist alongside glutamate, and contributes to the calcium influx that drives long-term potentiation, memory formation, and (when over-activated) excitotoxic cell death. See Neurotransmission and the NMDA Receptor for the full mechanism, the LTP-and-memory connection, the role in stroke and TBI excitotoxicity, and the clinically important NMDA-antagonist drug class.
- Urea cycle nitrogen donor and malate-aspartate shuttle carrier (L-aspartate) — the argininosuccinate synthetase (ASS1) step of the urea cycle is the only nitrogen-disposal pathway for one of the two nitrogens in each urea molecule, and aspartate is the obligatory donor. The malate-aspartate shuttle bridges cytosolic and mitochondrial NADH pools, supporting oxidative ATP synthesis in heart, liver, kidney, and brain. See Aspartate in the Urea Cycle for the full urea cycle anatomy, the five enzyme defects, and the management of acute and chronic hyperammonemia.
- Endocrine modulator via D-aspartate accumulation in steroidogenic tissues — the chiral D-isomer, generated by aspartate racemase, concentrates in the pituitary, testes, ovaries, and adrenal cortex, where it modulates GnRH, LH, FSH, growth hormone, testosterone, and progesterone. This is a completely separate biological role from L-aspartate's neurotransmitter and metabolic functions. See D-Aspartate and Testosterone for the tissue distribution, the HPG-axis pharmacology, and the controversial supplement market.
- TCA-cycle anaplerotic substrate and ATP-precursor donor (L-aspartate) — aspartate is in direct equilibrium with the TCA-cycle intermediate oxaloacetate via aspartate aminotransferase, supports gluconeogenesis during fasting, donates the N1 atom of every purine ring during de novo nucleotide synthesis (every ATP molecule), and participates in the purine nucleotide cycle that supports energy production in exercising muscle. See Aspartate and Energy Production for the metabolic networks and the history of aspartate as an ergogenic supplement.
The unifying picture is that aspartate sits at multiple strategic intersections in metabolism. It is both a neurotransmitter and a metabolic node; both a building block (for proteins, for purines) and a regulator (of urea synthesis, of endocrine signaling); both an L-form workhorse (across all four major roles) and a D-form specialist (in the endocrine system alone). The same amino acid, in two stereoisomeric forms, threads through nearly every major physiological system.
The therapeutic complication is that this same versatility makes targeted aspartate supplementation difficult to translate into reliable clinical benefit. The endogenous synthesis system (from oxaloacetate via AST) is robust under normal conditions, the blood-brain barrier restricts neural exposure, and feedback regulation in the HPG axis dampens the endocrine effects of exogenous D-aspartate. The supplement market for aspartate-containing products has historically promised more than the controlled trial evidence supports — with the partial exception of L-ornithine L-aspartate (LOLA) for hepatic encephalopathy, where the targeted clinical use is supported by modest controlled-trial evidence.
Research Papers: Neurotransmission & NMDA
- Bliss TV, Lømo T (1973). Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit — PubMed: Bliss-Lømo 1973 LTP
- Olney JW (1969). Brain lesions, obesity, and other disturbances in mice treated with monosodium glutamate — PubMed: Olney excitotoxicity 1969
- Tsien JZ et al. (1996). Hippocampal CA1 NMDA receptor and spatial memory — PubMed: Tsien 1996 CA1 knockout
- Choi DW (1988). Glutamate neurotoxicity and diseases of the nervous system — PubMed: Choi excitotoxicity review
- Bullock R et al. (1998). Excitatory amino acid release after severe human head injury — PubMed: Bullock 1998 TBI microdialysis
- Hardingham GE, Bading H (2010). Synaptic vs extrasynaptic NMDA receptor signalling — PubMed: Hardingham synaptic vs extrasynaptic
- Berman RM et al. (2000). Antidepressant effects of ketamine in depressed patients — PubMed: Berman 2000 ketamine antidepressant
- Reisberg B et al. (2003). Memantine in moderate-to-severe Alzheimer's disease — PubMed: Reisberg memantine Alzheimer's
- Rothstein JD et al. (2005). Beta-lactam antibiotics increase EAAT2 expression — PubMed: Rothstein 2005 EAAT2 ceftriaxone
- Hawkins RA (2009). The blood-brain barrier and glutamate — PubMed: Hawkins BBB and glutamate
Research Papers: Urea Cycle & Hyperammonemia
- Krebs HA, Henseleit K (1932). Original description of the urea cycle — PubMed: Krebs-Henseleit 1932
- Häberle J et al. (2019). Guidelines for urea cycle disorder diagnosis and management — PubMed: Häberle 2019 guidelines
- Brusilow SW et al. (1980). Sodium benzoate / phenylacetate for hyperammonemia — PubMed: Brusilow 1980 NEJM
- Batshaw ML et al. (2014). Longitudinal study of urea cycle disorders — PubMed: Batshaw longitudinal UCD
- Saheki T, Kobayashi K (2002). Citrin deficiency (mitochondrial aspartate-glutamate carrier) — PubMed: Saheki citrin deficiency
- Kido J et al. (2019). Long-term outcomes in urea cycle disorders — PubMed: Kido 2019 long-term UCD
- Kircher SG et al. (2017). L-ornithine L-aspartate (LOLA) meta-analysis for hepatic encephalopathy — PubMed: LOLA meta-analysis
- Vilstrup H et al. (2014). AASLD/EASL hepatic encephalopathy guideline — PubMed: Vilstrup 2014 HE guideline
- Bachmann C (2002). Mechanisms of hyperammonemia — PubMed: Bachmann hyperammonemia mechanism
- Wakabayashi Y et al. (1991). Argininosuccinate synthetase gene and chromosomal mapping — PubMed: ASS1 gene structure
Research Papers: D-Aspartate & Endocrine Function
- Topo E, Soricelli A, D'Aniello A et al. (2009). D-aspartic acid in the release and synthesis of LH and testosterone in humans and rats — PubMed: Topo D'Aniello 2009 RCT
- Willoughby DS, Leutholtz B (2013). D-aspartic acid + 28-day resistance training: no effect — PubMed: Willoughby 2013 failed replication
- Melville GW et al. (2015). 3 and 6 g/day D-aspartic acid in trained men — PubMed: Melville 2015 DAA dose
- Melville GW et al. (2017). 3-month DAA RCT — PubMed: Melville 2017 three-month RCT
- Roshanzamir F, Safavi SM (2017). D-aspartic acid and testosterone systematic review (negative) — PubMed: Roshanzamir systematic review
- D'Aniello A (2007). D-Aspartic acid: an endogenous amino acid with an important neuroendocrine role — PubMed: D'Aniello 2007 review
- D'Aniello A et al. (1996). D-aspartic acid in rat testosterone synthesis — PubMed: D'Aniello 1996 rat mechanism
- D'Aniello G et al. (2011). D-aspartate and sperm quality — PubMed: D-aspartate sperm quality
- Long Z et al. (2000). D-aspartate temporal/spatial distribution in pituitary and testis — PubMed: Long 2000 tissue distribution
- Furuchi T, Homma H (2005). Free D-aspartate in mammals — PubMed: Furuchi-Homma D-aspartate review
Research Papers: Energy Production & Mitochondrial Function
- Krebs HA, Johnson WA (1937). Original description of the citric acid cycle — PubMed: Krebs-Johnson 1937 TCA
- Owen OE et al. (2002). Anaplerosis and cataplerosis in TCA cycle function — PubMed: Owen 2002 anaplerosis
- Birsoy K et al. (2015). Mitochondrial ETC supports aspartate biosynthesis in proliferating cells — PubMed: Birsoy 2015 Cell
- Sullivan LB et al. (2015). Aspartate biosynthesis as essential function of respiration — PubMed: Sullivan 2015 Cell
- Lu M et al. (2010). The malate-aspartate shuttle — PubMed: Lu malate-aspartate review
- Tullson PC, Terjung RL (1991). Adenine nucleotide metabolism in contracting muscle — PubMed: Tullson-Terjung purine cycle
- Brand MD (2005). Efficiency and plasticity of mitochondrial energy transduction — PubMed: Brand 2005 mitochondrial efficiency
- Rosen H, Blumenthal A, McCallum J (1962). K-Mg aspartate on exercise tolerance — PubMed: Rosen 1962 K-Mg aspartate
- Wesson M et al. (1988). Aspartate and prolonged exercise — PubMed: Wesson aspartate exercise
- Cynober L (2002). Plasma amino acid levels and membrane transport — PubMed: Cynober plasma amino acids
Research Papers: Cross-Cutting (Mechanism, Status, Safety)
- Wolosker H et al. (2008). D-aspartate disposition in neuronal and endocrine tissues — PubMed: Wolosker D-aspartate
- Errico F et al. (2015). Free D-aspartate and synaptic plasticity — PubMed: Errico 2015 D-aspartate plasticity
- Wesson J et al. on aspartame metabolism safety reviews — PubMed: Aspartame safety reviews
- EFSA and FDA acceptable daily intake reviews for aspartame — PubMed: Aspartame ADI
- Vitamin B6 (pyridoxal-5-phosphate) and aminotransferase function — PubMed: B6 and aminotransferases
- Aspartate aminotransferase (AST) as clinical liver function marker — PubMed: AST as liver marker
- Plasma amino acid profiles in chronic disease — PubMed: Plasma amino acid profiles
- Branched-chain amino acid: aromatic amino acid ratio in cirrhosis — PubMed: BCAA:AAA ratio
- Aspartame and phenylketonuria contraindication — PubMed: Aspartame and PKU
- NIH Office of Dietary Supplements amino acid information — PubMed: Amino acid supplementation DRI
External Authoritative Resources
- Linus Pauling Institute — Micronutrient Information Center — broad amino acid coverage
- NIH Office of Dietary Supplements — Fact Sheets — comprehensive nutrient summaries
- UniProt — AST (mitochondrial aspartate aminotransferase) — protein database entry
- GeneReviews — Urea Cycle Disorders Overview — clinical genetics resource
- PubMed — All research on aspartic acid / aspartate (~200,000+ papers)
Connections
- Aspartic Acid (Main Page)
- Aspartic Acid for Neurotransmission & NMDA
- Aspartic Acid in the Urea Cycle
- D-Aspartate & Testosterone
- Aspartic Acid and Energy Production
- All Amino Acids
- Glutamic Acid
- Asparagine
- Arginine
- Glutamine
- Alanine
- Glycine
- Magnesium
- Zinc
- Potassium
- Vitamin B6
- Testosterone
- Liver Disease
- Stroke
- Fatigue
- Aspartame