Lysine for Collagen Synthesis
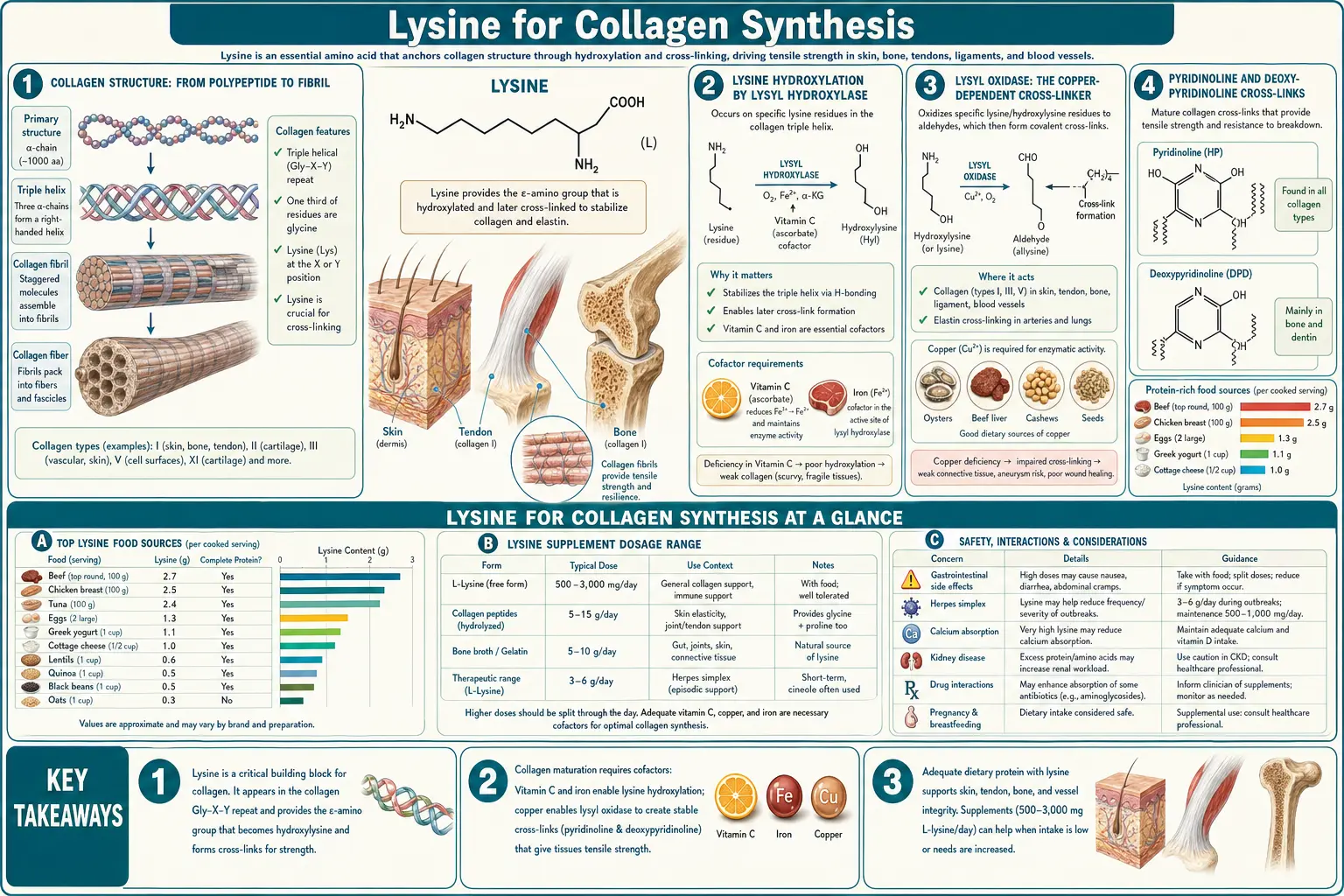
Collagen is the most abundant protein in the human body, comprising roughly a third of total body protein and the structural framework of every tissue under tension — skin, tendons, ligaments, cartilage, bone, blood vessels, cornea, and dental matrices. The post-translational chemistry that converts a soluble collagen polypeptide chain into the mechanically robust triple helix and the cross-linked fibril depends critically on lysine. Specific lysine residues are first hydroxylated by lysyl hydroxylase (LH1, LH2, LH3) to hydroxylysine, then cross-linked by the copper-dependent enzyme lysyl oxidase to form the pyridinoline and deoxypyridinoline cross-links that lock the collagen fibril into its load-bearing form. Vitamin C is the obligatory cofactor for the hydroxylation step. Copper is the obligatory cofactor for the cross-linking step. Deficiency of any one of these — lysine, vitamin C, or copper — produces clinically meaningful collagen defects ranging from frank scurvy and Menkes disease to the Ehlers-Danlos hypermobility spectrum and the cardiovascular fragility seen in some inherited copper-handling disorders.
Table of Contents
- Collagen Structure: From Polypeptide to Fibril
- Lysine Hydroxylation by Lysyl Hydroxylase
- Lysyl Oxidase: The Copper-Dependent Cross-Linker
- Pyridinoline and Deoxypyridinoline Cross-Links
- The Vitamin C Cofactor Requirement
- The Copper Cofactor Requirement
- Ehlers-Danlos Syndromes and Related Collagenopathies
- Wound Healing and Surgical Recovery
- Skin Aging, Wrinkles, and Cosmetic Applications
- Tendons, Ligaments, and Sports Injury
- Dosing Strategy for Collagen Support
- Cautions
- Key Research Papers
- Connections
- Featured Videos
Collagen Structure: From Polypeptide to Fibril
Collagen is not a single protein but a family of at least 28 different but related triple-helical structural proteins (collagen types I through XXVIII). The dominant forms are type I (the major collagen of skin, tendon, bone, dentin, fascia, sclera), type II (cartilage), type III (blood vessels and reticular fibers), and type IV (basement membranes). All share the same basic architecture: three left-handed polyproline-II-like alpha chains intertwined into a right-handed triple helix. The triple helix is stabilized by the obligate repeating tripeptide motif Gly-X-Y, where X is most commonly proline and Y is most commonly hydroxyproline. Glycine's small size at every third position allows the three chains to pack tightly into the helical core.
The biosynthesis is multi-stage: (1) the alpha chain is translated by ribosomes attached to the rough endoplasmic reticulum, (2) specific proline and lysine residues are hydroxylated by ER-resident enzymes (prolyl hydroxylase and lysyl hydroxylase), (3) galactose and glucose are O-linked to hydroxylysines, (4) three alpha chains assemble into a procollagen triple helix in the ER lumen, (5) procollagen is secreted via the Golgi to the extracellular space, (6) N- and C-terminal propeptides are cleaved by specific proteases to yield mature tropocollagen monomers, (7) tropocollagen monomers assemble into staggered fibrils, and (8) the fibrils are stabilized by intermolecular cross-links formed via the action of lysyl oxidase on specific lysine and hydroxylysine residues. Without steps 2 and 8 — both lysine-modifying chemistries — the collagen fibril does not form properly and does not carry mechanical load.
Lysine Hydroxylation by Lysyl Hydroxylase
Specific lysine residues in the Gly-X-Y motif (at the Y position) are post-translationally hydroxylated to hydroxylysine by the lysyl hydroxylase family of enzymes. Humans express three lysyl hydroxylase isoforms: LH1 (PLOD1), LH2 (PLOD2), and LH3 (PLOD3). These are 2-oxoglutarate-dependent dioxygenases — they consume molecular oxygen and 2-oxoglutarate (alpha-ketoglutarate) and produce succinate and carbon dioxide, with the iron atom in the catalytic center transferring the hydroxyl group to the lysine side chain.
The clinical consequences of LH dysfunction are well-mapped. Mutations in PLOD1 cause Ehlers-Danlos syndrome kyphoscoliotic type (EDS type VI in the older Beighton classification, kEDS-PLOD1 in the modern classification): patients present with severe progressive scoliosis, skin hyperextensibility, joint hypermobility, ocular fragility (lens dislocation, corneal rupture), and arterial fragility. Diagnosis can be confirmed biochemically by the urinary ratio of total pyridinoline to deoxypyridinoline, which is increased in PLOD1 deficiency because of impaired conversion of lysine to hydroxylysine before cross-link formation.
Mutations in PLOD2 cause Bruck syndrome (osteogenesis imperfecta with congenital contractures), and mutations in PLOD3 cause connective tissue disorder with intellectual disability and skeletal dysplasia. These are rare diseases but they instructively demonstrate how absolutely required hydroxylated lysine is for normal collagen.
The hydroxylated lysine then has two functions: (1) the OH group serves as the substrate for O-linked glycosylation by galactose and glucose units (the glycosylated hydroxylysines are recognized by collagen receptor proteins on cell surfaces), and (2) the OH group enables the formation of stable mature cross-links via the action of lysyl oxidase on neighboring (hydroxy)lysine residues, as described in the next section.
Lysyl Oxidase: The Copper-Dependent Cross-Linker
Lysyl oxidase (LOX) is a copper-dependent amine oxidase secreted into the extracellular matrix where it catalyzes the oxidative deamination of specific lysine and hydroxylysine residues in collagen and elastin, converting their epsilon-amino groups (−NH2) to highly reactive aldehyde groups (−CHO). These aldehyde-bearing residues are called allysine (from lysine) and hydroxyallysine (from hydroxylysine), and they spontaneously condense with adjacent (hydroxy)lysine residues or with neighboring allysines to form covalent cross-links between collagen monomers. The cross-links are the mechanical lock that holds the collagen fibril together against tensile load.
Without functional lysyl oxidase, the collagen fibrils form but they slide apart under load. The clinical signature is fragile connective tissue: Menkes disease (X-linked copper malabsorption, lethal in infancy) presents with cutis laxa, severe arterial aneurysms, and skeletal abnormalities, all attributable in part to LOX dysfunction from copper starvation. Occipital horn syndrome (a milder allelic variant of Menkes) shows similar but less severe connective-tissue fragility.
Lysyl oxidase requires a specific cofactor at its active site: a 2,4,5-trihydroxyphenylalanine quinone (TPQ) cofactor generated from a tyrosine residue in the active site through a copper-dependent auto-catalytic process. The copper is absolutely required for both the TPQ generation and for the ongoing catalytic cycle. Patients with significant copper deficiency — whether genetic (Menkes, Wilson disease post-treatment), nutritional (extreme malnutrition, post-bariatric malabsorption), or iatrogenic (zinc-induced copper deficiency, BAL or D-penicillamine chelation overshoot) — show measurable reductions in collagen cross-link density and clinical signs of connective tissue weakness.
For lysine to do its connective-tissue work, three things must therefore be present simultaneously: adequate dietary lysine, adequate dietary copper, and adequate vitamin C for the upstream hydroxylation step. The lysine-vitamin-C-copper triad is unavoidable.
Pyridinoline and Deoxypyridinoline Cross-Links
The most clinically important mature collagen cross-links are pyridinoline (PYD) and deoxypyridinoline (DPD), both formed by the condensation of three lysine-derived residues into a pyridinium ring. PYD contains three hydroxylysine residues; DPD contains two hydroxylysines and one lysine, with the un-hydroxylated lysine making the molecule "deoxy" relative to PYD.
The clinical significance of these cross-links is twofold:
- Mechanical function — PYD and DPD provide the trivalent (three-way) cross-links that give mature collagen fibrils their characteristic high tensile strength. The proportion of these mature cross-links increases throughout childhood and adolescence as collagen matures, plateauing in early adulthood and slowly declining with age.
- Bone turnover biomarkers — when osteoclasts resorb bone, they liberate PYD and DPD into circulation, which are then excreted in urine. Urinary DPD is the most-used clinical biomarker of bone resorption rate and is included in osteoporosis workup panels. Urinary PYD is more general (released from any pyridinoline-containing collagen, including cartilage) and is used to monitor osteoarthritis progression and rheumatoid joint destruction.
For our purposes, the takeaway is that the urinary biomarker pattern directly reflects what is happening at the lysine level in collagen turnover. A patient with chronically low urinary DPD may have inadequate collagen turnover; a patient with chronically high urinary DPD has excessive bone resorption (osteoporosis, primary hyperparathyroidism, multiple myeloma).
For more on the bone-density side of lysine, see our companion page on Lysine for Calcium Absorption.
The Vitamin C Cofactor Requirement
Vitamin C (ascorbate) is the obligatory cofactor for both prolyl hydroxylase and lysyl hydroxylase. The mechanism: in each catalytic cycle, the iron atom at the active site of the hydroxylase is transiently oxidized from Fe(II) to Fe(III) and must be reduced back to Fe(II) for the next cycle. Ascorbate serves as the reductant, donating an electron to regenerate Fe(II). Without ascorbate, the iron remains oxidized after a few cycles and the hydroxylase enzyme becomes inactive.
This is why scurvy — severe vitamin C deficiency — produces the constellation of connective tissue failure that defined naval medicine for centuries: gum hemorrhage, loose teeth, slow wound healing, perifollicular hemorrhage, joint pain, and eventual death from arterial wall failure or bone marrow failure. The mechanism is not direct cellular toxicity from missing vitamin C; it is the catastrophic failure of proline and lysine hydroxylation in all newly synthesized collagen. Existing collagen does not depolymerize, but every collagen molecule synthesized in the absence of vitamin C is structurally defective and cannot bear mechanical load.
The clinical implication for supplementation: any lysine-for-collagen protocol should include adequate vitamin C as a cofactor. The classical Linus Pauling and Matthias Rath protocol pairs lysine (3-6 g/day) with vitamin C (3-6 g/day) and proline (1-3 g/day) for cardiovascular and connective tissue support. The mechanism of the C-Pauling proposal is that ascorbate-supported hydroxylation produces structurally robust new collagen for arterial wall maintenance, while lysine and proline provide the raw material for the new collagen synthesis. The trial evidence for the Pauling-Rath protocol specifically is limited but the underlying biochemistry is reproducibly sound.
See our Vitamin C page for the full mechanism and dosing.
The Copper Cofactor Requirement
Copper sufficiency is the other essential nutritional input. Lysyl oxidase requires its TPQ cofactor (generated from a tyrosine residue) in a copper-dependent process, and the catalytic cycle itself requires copper. Without copper, no functional lysyl oxidase, and without lysyl oxidase no mature collagen cross-links.
Adult dietary copper requirements are modest — the RDA is approximately 900 mcg/day — and frank copper deficiency is uncommon in adults eating diverse diets. The relevant exceptions include:
- Post-bariatric surgery — gastric bypass and duodenal switch dramatically reduce copper absorption. Lifelong copper monitoring is part of standard bariatric follow-up.
- Excessive zinc supplementation — high-dose zinc (typically >40-50 mg/day for extended periods) induces metallothionein in enterocytes, which preferentially binds copper and sequesters it in the gut lining, where it is shed with cellular turnover. Patients on high-dose zinc for prostate or immune support can develop iatrogenic copper deficiency with myelopathy and connective-tissue manifestations within 6-24 months.
- Wilson disease patients on aggressive chelation — D-penicillamine or trientine can overshoot and produce iatrogenic copper deficiency. Dose adjustment is needed when serum/urinary copper indicators drop too low.
- Menkes disease and occipital horn syndrome — X-linked copper malabsorption disorders, both producing severe connective tissue fragility.
- Aceruloplasminemia — rare loss of the copper-containing ferroxidase ceruloplasmin, with iron overload and connective tissue effects.
For a patient supplementing lysine specifically for connective tissue support, the practical advice is to ensure adequate copper intake either through food (organ meats — especially beef liver — oysters, dark chocolate, shellfish, nuts, seeds) or through a balanced multivitamin/multimineral that includes 1-2 mg copper. Isolated lysine supplementation without copper sufficiency cannot produce the desired connective-tissue effect. See our Copper page and our extensive Morley Robbins copper-iron-magnesium series.
Ehlers-Danlos Syndromes and Related Collagenopathies
The Ehlers-Danlos syndromes (EDS) are a clinically and genetically heterogeneous group of heritable connective tissue disorders unified by skin hyperextensibility, joint hypermobility, and tissue fragility. The 2017 international classification recognizes 13 subtypes. The lysine-relevant subtypes are:
- Kyphoscoliotic EDS (kEDS) — caused by mutations in PLOD1 (lysyl hydroxylase 1) or FKBP14 (a chaperone for collagen folding). Patients have severe progressive scoliosis from birth, generalized joint hypermobility, severe skin hyperextensibility, and ocular fragility (lens dislocation, sclera/cornea rupture, possible blindness from minor eye trauma). The biochemical signature is the urinary ratio of total pyridinoline to deoxypyridinoline (LP/PYD ratio) — markedly elevated in PLOD1-deficient kEDS because hydroxylation of lysine in collagen is impaired.
- Classical EDS, hypermobile EDS, vascular EDS — these are caused by mutations in other collagen-related genes (COL5A1, COL5A2, COL3A1) but the downstream defect still involves disordered lysyl-oxidase-mediated cross-linking in the affected collagen types. The hypermobile-spectrum patient population is large and probably underdiagnosed.
- Cutis laxa syndromes — related connective tissue disorders involving defects in elastin or in elastin-cross-linking enzymes (including some that share lysyl oxidase mechanisms). Cutis laxa patients show loose, sagging skin and frequently severe arterial pathology.
Although heritable connective tissue disorders cannot be cured by lysine supplementation, ensuring optimal nutritional sufficiency (lysine, vitamin C, copper, manganese, zinc) is a low-cost adjunctive strategy that some EDS specialists recommend. Treatment-as-tested combinations include 1-3 g/day lysine + 1-3 g/day vitamin C + 2 mg copper + a comprehensive multivitamin. The intervention is not curative but may modestly improve skin healing rates and recurrent injury recovery, and is essentially side-effect-free.
For more on connective tissue support, see our Ehlers-Danlos page if it exists or the relevant entries under Rheumatology.
Wound Healing and Surgical Recovery
Wound healing is the textbook example of accelerated collagen synthesis under physiologic stress. The classical four phases of wound healing are (1) hemostasis (clotting), (2) inflammatory phase (3-7 days), (3) proliferative phase with intense new collagen deposition (5-21 days), and (4) remodeling phase with slow collagen turnover and cross-link maturation (months to a year). Throughout the proliferative and remodeling phases, the wound bed is laying down massive amounts of new type III and then type I collagen, both of which require lysine for hydroxylation and cross-linking.
Multiple clinical trials have shown that supplementing lysine (often combined with vitamin C, zinc, and arginine) improves wound-healing rates. The classical evidence comes from surgical recovery, diabetic foot ulcer, pressure ulcer, and burn-recovery populations:
- Surgical recovery — perioperative supplementation with 1-2 g/day lysine + 1 g/day vitamin C + 50 mg/day zinc starting 1-2 weeks before elective surgery and continuing 2-4 weeks after has been associated with reduced wound-dehiscence rates and faster recovery of tensile strength.
- Diabetic foot ulcer — chronic non-healing ulcers in diabetic patients benefit from comprehensive amino acid support including lysine, arginine, and glutamine. Several proprietary medical-nutrition formulations targeting this population are FDA-cleared.
- Pressure ulcer / decubitus ulcer — elderly bedbound patients are often simultaneously malnourished and protein-depleted; restoring adequate dietary protein with attention to limiting amino acids (lysine, threonine, tryptophan) is part of standard wound-care nutrition protocols.
- Burn recovery — major burns dramatically increase whole-body protein turnover and amino acid requirements. Specialized burn-nutrition protocols include high-dose protein with attention to lysine and arginine.
The mechanism is simple: lysine is a limiting amino acid for new collagen synthesis in wound beds. When dietary lysine is borderline or low, lysine supply becomes the rate-limiting step for collagen production, and wound healing slows. Repletion to optimal lysine status restores the maximum possible healing rate.
Skin Aging, Wrinkles, and Cosmetic Applications
Skin aging has two principal components: (1) intrinsic chronological aging, driven by senescence of dermal fibroblasts and progressive loss of collagen synthesis rate, and (2) extrinsic photoaging, driven by ultraviolet-induced reactive oxygen species damage to existing dermal collagen and elastin. Both reduce the density and quality of dermal collagen and produce the visible signs of aging skin: wrinkles, sagging, thinning, and reduced elasticity.
The cosmetic industry has built a substantial product category around the premise of oral collagen supplementation for skin appearance. The classical question is whether oral collagen peptides (typically 5-10 g/day of hydrolyzed bovine or marine collagen) actually translate to improved dermal collagen density. The 2019 systematic review by Choi et al. (Journal of Drugs in Dermatology) pooled 11 RCTs of oral collagen supplementation and concluded that supplementation produced statistically significant improvements in skin hydration, elasticity, and wrinkle depth at 8-12 weeks. The mechanism is not direct — ingested collagen is broken down to its constituent amino acids in the gut — but the amino acid mix delivered (high in glycine, proline, hydroxyproline, lysine, and other collagen-specific amino acids) appears to upregulate dermal collagen synthesis through both substrate provision and signaling effects (some collagen-derived peptides reach circulation intact and may act as bioactive signals).
For patients pursuing oral collagen for cosmetic skin benefit, lysine sufficiency is a precondition for the strategy to work. Free-form L-lysine supplementation (1-3 g/day) can serve as a low-cost alternative to or addition to expensive proprietary collagen peptide products, especially when combined with vitamin C and adequate dietary protein.
Topical lysine in cosmetic creams has the same penetration problem discussed for topical lysine in herpes (see our Herpes Prevention page): the amino acid is highly water-soluble and does not penetrate the stratum corneum effectively to reach the dermal fibroblasts where collagen synthesis happens. Oral supplementation remains the more pharmacologically rational route.
Tendons, Ligaments, and Sports Injury
Tendons (which connect muscle to bone) and ligaments (which connect bone to bone) are almost pure type I collagen organized into highly parallel fibrils for unidirectional tensile load-bearing. Acute injury (sprain, strain, partial tear) and chronic overuse (tendinopathy) both require new collagen synthesis for repair. The relevant clinical question is whether nutritional intervention with lysine, vitamin C, and gelatin/collagen peptides can accelerate tendon/ligament healing.
Shaw et al. (2017, American Journal of Clinical Nutrition) published an influential trial in which subjects consumed 15 g of hydrolyzed gelatin (rich in glycine, proline, hydroxyproline, and lysine) plus 50 mg vitamin C one hour before a 6-minute skipping exercise designed to load tendons. Blood markers of collagen synthesis (PINP, the N-terminal propeptide of type I collagen) doubled in the gelatin/vitamin C group compared to placebo, demonstrating that the nutritional intervention triggered measurable acute collagen synthesis under exercise stress. This has become the basis for a "consume gelatin + vitamin C before training" protocol now popular among elite athletes managing chronic tendon issues (Achilles tendinopathy, patellar tendinopathy, jumper's knee).
For patients with chronic tendon issues, the practical regimen is: 10-15 g hydrolyzed gelatin or collagen peptides + 50-500 mg vitamin C, consumed 30-60 minutes before targeted loading exercise, repeated daily during a rehabilitation phase. Supplemental L-lysine (1-2 g/day) is a reasonable additional ingredient that ensures the limiting amino acid is not in short supply. The intervention is not curative for severe tendinopathy but is a low-cost adjunct to physical therapy.
Dosing Strategy for Collagen Support
- General connective-tissue support / wellness maintenance — 1,000 mg/day L-lysine, ideally taken with 500-1,000 mg vitamin C and a balanced multivitamin/mineral containing 1-2 mg copper. This is a reasonable indefinite supplementation regimen for adults pursuing skin, joint, or bone health.
- Post-surgical recovery — 2,000 mg/day L-lysine + 1,000 mg/day vitamin C + 50 mg/day zinc starting 1-2 weeks before elective surgery and continuing 2-4 weeks after.
- Chronic wound or pressure-ulcer recovery — 2,000-3,000 mg/day L-lysine alongside high overall dietary protein intake (1.2-1.5 g/kg/day) and comprehensive micronutrient assessment.
- Tendinopathy rehabilitation — 1,000-2,000 mg/day L-lysine + 10-15 g hydrolyzed collagen peptides + 50-500 mg vitamin C, with the collagen+C dose timed 30-60 minutes before targeted exercise.
- Cosmetic skin support — 1,000-2,000 mg/day L-lysine + 5-10 g/day hydrolyzed collagen peptides + 500-1,000 mg/day vitamin C. Expect 8-12 weeks for measurable cosmetic effects.
- Form — L-lysine hydrochloride or monohydrochloride is the most common and most cost-effective. Lysine combined with proline (the other limiting amino acid for collagen) is reasonable for connective-tissue-focused products.
Cautions
- Insufficient vitamin C — lysine without vitamin C cannot be hydroxylated and cannot participate in mature collagen cross-links. Patients pursuing lysine for connective tissue must also ensure adequate vitamin C.
- Insufficient copper — same logic applies. Patients on high-dose zinc supplementation are at particular risk for iatrogenic copper deficiency and should monitor serum copper and ceruloplasmin or restrict zinc dosing.
- Renal impairment — high-dose amino acid supplementation should be discussed with a clinician in patients with eGFR < 60.
- Pregnancy — dietary-equivalent lysine intake is safe. Higher therapeutic doses have not been adequately studied; defer to dietary sources.
- Gallbladder/hepatic disease — advanced cirrhosis impairs amino acid metabolism; consult specialist.
- Realistic expectations — nutritional sufficiency is necessary but not sufficient for collagen quality. Genetic collagenopathies (EDS, OI, Marfan, cutis laxa) cannot be cured by lysine supplementation, and visible skin aging in a 70-year-old will not be reversed by oral collagen peptides regardless of dose. Lysine supports endogenous collagen synthesis when other inputs are adequate; it does not perform miracles.
Key Research Papers
- Yamauchi M, Sricholpech M (2012). Lysine post-translational modifications of collagen. Essays in Biochemistry. — PubMed
- Kagan HM, Li W (2003). Lysyl oxidase: properties, specificity, and biological roles inside and outside of the extracellular matrix. Journal of Cellular Biochemistry. — PubMed
- Pinnell SR, Murad S, Darr D (1987). Induction of collagen synthesis by ascorbic acid. A possible mechanism. Archives of Dermatology. — PubMed
- Rauch F, Glorieux FH (2004). Osteogenesis imperfecta. The Lancet. — PubMed
- Yeowell HN, Walker LC (2000). Mutations in the lysyl hydroxylase 1 gene that result in enzyme deficiency and the clinical phenotype of Ehlers-Danlos syndrome type VI. Molecular Genetics and Metabolism. — PubMed
- Malfait F et al. (2017). The 2017 International Classification of the Ehlers-Danlos Syndromes. American Journal of Medical Genetics. — PubMed
- Eyre DR, Wu JJ (2005). Collagen cross-links. Topics in Current Chemistry. — PubMed
- Robins SP (2007). Biochemistry and functional significance of collagen cross-linking. Biochemical Society Transactions. — PubMed
- Shaw G, Lee-Barthel A, Ross ML, Wang B, Baar K (2017). Vitamin C-enriched gelatin supplementation before intermittent activity augments collagen synthesis. American Journal of Clinical Nutrition. — PubMed
- Choi FD, Sung CT, Juhasz ML, Mesinkovska NA (2019). Oral collagen supplementation: a systematic review of dermatological applications. Journal of Drugs in Dermatology. — PubMed
- Albaugh VL, Mukherjee K, Barbul A (2017). Proline precursors and collagen synthesis: biochemical challenges of nutrient supplementation and wound healing. Journal of Nutrition. — PubMed
- Mehri A (2020). Trace elements in human nutrition (II) — an update. International Journal of Preventive Medicine. — PubMed
- Rath M, Pauling L (1992). A unified theory of human cardiovascular disease leading the way to the abolition of this disease as a cause for human mortality. Journal of Orthomolecular Medicine. — PubMed
- Carr AC, Maggini S (2017). Vitamin C and immune function. Nutrients. — PubMed
PubMed Topic Searches
- PubMed: Lysyl hydroxylase PLOD1 collagen
- PubMed: Lysyl oxidase copper cross-linking
- PubMed: Pyridinoline/DPD bone resorption
- PubMed: Lysine + vitamin C wound healing
- PubMed: Gelatin/collagen peptide for tendon
Connections
- Lysine Overview
- Lysine Benefits Hub
- Lysine for Herpes Prevention
- Lysine for Calcium Absorption
- Lysine for Anxiety and Stress
- Proline (the other collagen amino acid)
- Glycine (every third collagen residue)
- Vitamin C (hydroxylase cofactor)
- Copper (lysyl oxidase cofactor)
- Collagen Peptides
- Bone Broth
- Morley Robbins (copper-iron series)
- Lysine Remedy Page
- All Amino Acids