Inositol: History and Discovery
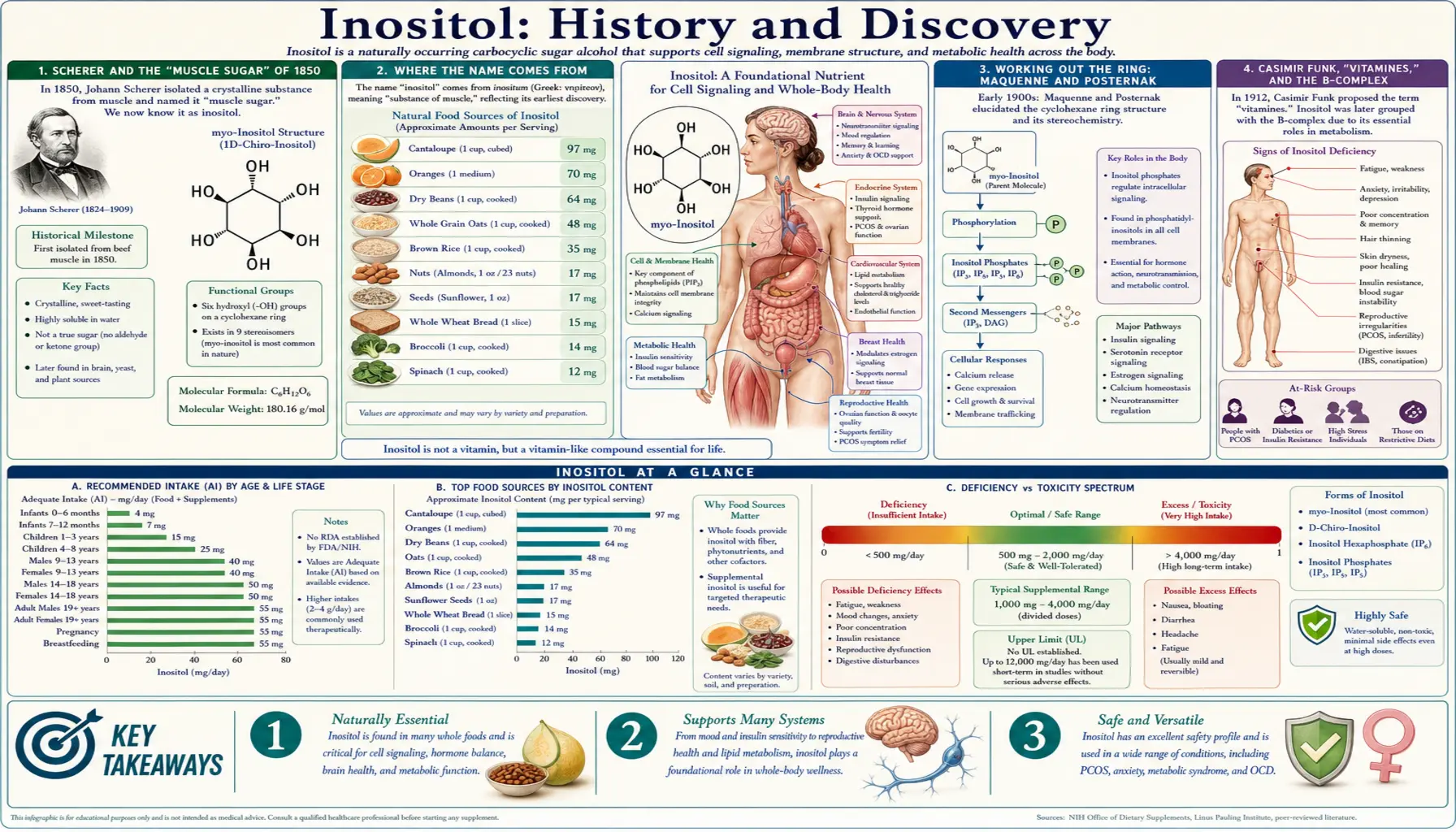
In 1850 a young German physician-chemist named Johann Joseph Scherer boiled down an extract of muscle, watched sweet crystals separate out, and gave the new substance a name built from the Greek word for muscle: inosit. He had discovered what we now call inositol. The story that follows is unusually long and unusually honest about its own twists — a compound first taken for a kind of "muscle sugar," later crowned as vitamin B8, then quietly stripped of that title when scientists realised the human body makes its own. Along the way real, named people worked out its ring-shaped structure, mapped its mirror-image forms, traced it into the membranes of every living cell, and finally — in a discovery that won the Lasker, Wolf, and Shaw prizes but never a Nobel — showed that a fragment of inositol is one of the body's master chemical messengers. Where the record is firm we say so; where a date or a name is uncertain or disputed, we mark it as such, and we do not invent a discoverer where the evidence names none.
Table of Contents
- Scherer and the "Muscle Sugar" of 1850
- Where the Name Comes From
- Working Out the Ring: Maquenne and Posternak
- Casimir Funk, "Vitamines," and the B-Complex
- The Brief Life of Vitamin B8
- Why Inositol Lost Its Vitamin Status
- From a Curiosity to the Heart of the Cell
- Berridge and the Second-Messenger Revolution
- The Modern Revival
- Research Papers and References
- Connections
- Featured Videos
Scherer and the "Muscle Sugar" of 1850
The discovery of inositol belongs to Johann Joseph Scherer (1814–1869), a German physician and chemist remembered as one of the founders of clinical and physiological chemistry. In 1850 Scherer isolated a previously unknown crystalline substance from the extract of mammalian muscle. To his eye it behaved much like a sugar — it was sweet, it crystallised cleanly — and because it had come out of muscle tissue it was sometimes described in the early literature as a kind of "muscle sugar." That first impression was understandable but, as later chemistry would show, misleading: inositol shares the bare molecular formula of glucose (C6H12O6) yet is built on a completely different skeleton, a six-membered carbon ring rather than the open-chain or pyranose form of an ordinary sugar.
It is worth being precise about what Scherer did and did not do. He isolated and named the compound; he did not yet know its true structure, which would take another generation of chemists to settle (see below). What matters for the history is that 1850 is the firm, repeatedly documented starting point — the moment a new natural compound entered the scientific record — and that it is tied to a single, named discoverer working with muscle tissue. Nearly everything else in inositol's long story unfolds from that one extraction.
Where the Name Comes From
The name inositol is a small fossil of its discovery. Scherer built it from the ancient Greek ὶς (transliterated is / inos), meaning "sinew" or "fibre" — a direct nod to the muscle tissue the compound first came out of. To that root he added the chemical ending that marked it, in the language of the day, as a sugar-like substance; the modern English form inositol carries the "-ol" ending used for alcohols, reflecting that inositol is technically a sugar alcohol (a polyol), with a hydroxyl group on every one of its six ring carbons.
The most familiar form of the molecule is myo-inositol, and that prefix has its own etymological echo: myo- is from the Greek for muscle as well, the same root that gives us words like myology. So the dominant biological isomer of inositol is, quite literally, named "muscle inositol" twice over. Other isomers carry descriptive Greek prefixes too — scyllo-, chiro- ("hand," signalling that those forms are chiral, or handed), neo-, epi-, and so on — a naming scheme that was later regularised into a formal system of cyclitol nomenclature. The thread running through all of it is the original 1850 observation: a sweet, ring-shaped alcohol first pulled from muscle.
Working Out the Ring: Maquenne and Posternak
Knowing that a compound exists is not the same as knowing what it looks like, and inositol's true architecture took decades to pin down. A major step came in 1887, when the French chemist Léon Maquenne purified inositol (notably from plant leaves as well as animal tissue) and established that it was a cyclohexanehexol — a six-carbon ring (a cyclohexane) carrying six hydroxyl groups, one on each carbon. This was the insight that finally separated inositol conceptually from the ordinary sugars it had been lumped with: it was not a sugar at all in the structural sense, but a ringed polyalcohol.
That ring, however, can be assembled in several distinct three-dimensional ways. Because each of the six hydroxyl groups can point "up" or "down" relative to the ring, inositol exists as a family of nine stereoisomers — same atoms, different spatial arrangements. Untangling which arrangement corresponded to the inositol found in living tissue was the achievement most associated with the Swiss chemist Théodore Posternak, whose careful work in the 1930s and 1940s established the configuration of the principal natural form, myo-inositol, and who helped develop the logical, self-consistent system for naming the various cyclitols that chemists still draw on today. (Some early reports pair Posternak's name with that of G. Dangschat in this structural work; we name Posternak here as the figure most consistently credited, without overstating the precise division of labour.) By the mid-twentieth century, then, a compound discovered as a mysterious muscle crystal in 1850 had a fully worked-out structure and a rational family tree of isomers.
Casimir Funk, "Vitamines," and the B-Complex
To understand how inositol briefly became a "vitamin" at all, it helps to step back to the idea that made vitamins thinkable. In 1912 the Polish-born biochemist Casimir Funk (1884–1967), then working in London, coined the word "vitamine." He had been studying beriberi — the nerve-and-heart disease that struck populations living on polished white rice — and was convinced that such diseases were caused by the absence of tiny, essential dietary factors rather than by a poison or an infection. Believing (incorrectly, as it turned out) that these factors were all chemical amines vital to life, he fused vita ("life") with amine to make vitamine; the trailing "e" was later dropped once it became clear that not all of them are amines, leaving the modern vitamin. Funk laid out the concept in his 1912 paper "The Etiology of the Deficiency Diseases," proposing that distinct factors prevented beriberi, scurvy, pellagra, and rickets.
Funk's framework opened a decades-long hunt for these "accessory food factors," and the water-soluble ones were gathered under the umbrella of the vitamin B complex and assigned numbers — B1 (thiamine), B2 (riboflavin), and onward. The numbering grew crowded and, in places, provisional: several substances were given B-numbers on early, animal-based evidence and were later demoted when they turned out not to meet the strict definition of a vitamin. Inositol was one of those provisional members, and it is to its short career as vitamin B8 that we now turn.
The Brief Life of Vitamin B8
Inositol's nutritional moment arrived around 1940, when researchers studying the diets of laboratory animals reported that it behaved like an essential growth factor. The most evocative early finding was that mice deprived of inositol developed alopecia — they lost their fur — and that adding inositol back to the diet restored healthy hair and skin. This gave inositol its memorable early nickname, the "mouse antialopecia factor," and similar deficiency signs (sometimes described as "spectacle eyes" in rats) reinforced the impression that it was a genuine dietary essential. Work in this period — including studies by D. W. Woolley on inositol's biological role and on the alopecia it could reverse, and contemporary notes in Science on the antialopecia factor — cemented inositol's place, for a time, among the B-complex vitamins under the label vitamin B8.
It is fair to call this an honest mistake rather than a fabrication. The animal evidence was real: under those particular experimental diets, inositol really was required for normal growth and coat. The error lay in generalising from rodents on artificial diets to the everyday human, and in assuming that "essential in the diet of a deprived mouse" meant "essential in the diet of a person." As we will see, that assumption did not survive contact with later human biochemistry.
Why Inositol Lost Its Vitamin Status
The strict definition of a vitamin is demanding: a substance must be required for normal physiology yet not made by the body in adequate amounts, so that it has to be supplied by the diet. Inositol fails the second half of that test. The human body synthesises its own inositol, principally in the kidneys, where the enzyme inositol-1-phosphate synthase builds it from glucose-6-phosphate; the kidneys alone are estimated to produce on the order of a couple of grams of myo-inositol per day, supplemented by what we absorb from food. Because a healthy person manufactures inositol in quantity, it cannot, by definition, be a true dietary vitamin.
For this reason inositol was reclassified out of the vitamin category and is now usually described as a pseudovitamin or vitamin-like compound — the same fate, for the same reason, that befell several other one-time B-numbers. This is exactly the kind of "former vitamin" story worth flagging plainly: vitamin B8 is a historical label, not a current scientific classification. None of this diminishes inositol's biological importance. It simply relocates it — from the short list of substances we must eat to survive, to the much larger and more interesting category of molecules the body makes and uses for itself. And as the next two sections show, what the body uses inositol for turned out to be far more central than anyone studying hairless mice in 1940 could have guessed.
From a Curiosity to the Heart of the Cell
Through the first half of the twentieth century, inositol was mostly a chemical curiosity with a fading vitamin reputation. Its scientific fortunes changed once researchers found it embedded in the fatty membranes of living cells. In 1949, the biochemist Jordi Folch-Pi, working on brain tissue, identified inositol-containing phospholipids — the family we now call the phosphoinositides, built around a core lipid named phosphatidylinositol. Inositol, in other words, was not just floating free in muscle and citrus fruit; a piece of it was wired into the very fabric of cell membranes.
The next clue came from an experiment that seemed, at first, almost incidental. In 1953, the husband-and-wife team Lowell and Mabel Hokin were studying pancreas tissue and noticed that when they stimulated the cells with the neurotransmitter acetylcholine, the cells dramatically ramped up the turnover of their inositol-containing lipids — soaking up radioactively labelled phosphate into phosphatidylinositol. This "phosphoinositide effect" was a genuine puzzle: why should poking a cell with a hormone-like signal cause it to churn through a minor membrane lipid? For roughly three decades the observation sat in the literature as a striking but unexplained phenomenon. The answer, when it finally came, would rewrite the textbooks of cell biology.
Berridge and the Second-Messenger Revolution
The breakthrough belongs above all to the British physiologist Sir Michael J. Berridge (1938–2020) and his collaborators in the early 1980s. Berridge realised that the long-mysterious phosphoinositide effect was the visible footprint of a signalling system. When a hormone or neurotransmitter docks onto a receptor on the cell surface, an enzyme splits the membrane lipid PIP2 (a doubly-phosphorylated phosphatidylinositol) into two pieces — and one of those pieces, a small water-soluble fragment of inositol called inositol 1,4,5-trisphosphate (IP3), races into the cell interior and triggers the release of stored calcium. Calcium, in turn, sets off the cell's response. IP3 was, in the language of the field, a second messenger: the relay that carries a signal from the cell's outer membrane to its inner machinery.
The key experimental proof was published in 1983 in Nature by H. Streb, R. F. Irvine, M. J. Berridge, and I. Schulz, who showed directly that IP3 releases calcium from an internal store in pancreatic cells. The following year Berridge and Irvine drew the threads together in an influential Nature review framing IP3 as a general second messenger in cellular signal transduction. The discovery transformed inositol from a half-forgotten ex-vitamin into a molecule sitting at the centre of how virtually every cell in the body hears and obeys its hormones — insulin, serotonin, and many others all lean on this same phosphoinositide relay.
This work earned Berridge some of science's highest honours: the Albert Lasker Basic Medical Research Award (1989), the Wolf Prize in Medicine (shared with Yasutomi Nishizuka, 1994–95), and the inaugural Shaw Prize in Life Science and Medicine (2005). It is worth stating clearly, because the claim is easy to garble: Berridge did not receive a Nobel Prize, and there is no Nobel Prize awarded specifically for inositol. The honest summary is that inositol's pivotal scientific moment was Lasker-, Wolf-, and Shaw-recognised work on cell signalling — not a Nobel, and we do not claim one.
The Modern Revival
The discovery that IP3 is a master second messenger reframed inositol entirely. If inositol-based signalling underpins how cells respond to insulin, to follicle-stimulating hormone, to thyroid-stimulating hormone, and to serotonin, then problems with inositol availability or recycling might plausibly contribute to conditions spanning metabolism, reproduction, and mood. From the 1990s onward, that logic drove a wave of clinical research — most prominently into polycystic ovary syndrome (PCOS), where myo-inositol and its sibling D-chiro-inositol act as messengers of insulin signalling, and into anxiety, panic disorder, and depression, where the link runs through serotonin's use of the same pathway. A widely cited 2014 review even bears the apt title "Inositol: history of an effective therapy for Polycystic Ovary Syndrome," capturing how a nineteenth-century muscle extract became a twenty-first-century supplement.
Two honest cautions close the history. First, the arc from "vitamin B8" to "pseudovitamin" is a reminder that scientific classification is provisional: a substance can be promoted on partial evidence and demoted on fuller evidence without ever changing what it actually does in the body. Second, the modern enthusiasm for inositol supplements, while grounded in real signalling biology and a growing clinical literature, is still an active research area rather than settled fact — the strength of the evidence varies a good deal from one condition to the next. The detailed clinical picture, dosing, and cautions are covered in the companion Inositol Benefits articles and on the main Inositol page; this article has been concerned with how the molecule was found, named, mis-classified, re-understood, and finally placed at the centre of cell biology.
Research Papers and References
The list below combines peer-reviewed sources on inositol's biochemistry and clinical history with curated PubMed topic-search links and authoritative reference pages. Historical milestones discussed in the article — Scherer's 1850 isolation, Maquenne's 1887 structural work, Posternak's 1930s–40s stereochemistry, the 1940-era "mouse antialopecia factor," Folch's 1949 phospholipid work, and the Hokins' 1953 phosphoinositide effect — are named in the text as historical sources; their original publications predate stable digital identifiers and so are not hyperlinked here. Author names, titles, and journals are given as plain text; only a stable DOI, PMID, or archive link is hyperlinked, and each opens in a new tab.
- Streb H, Irvine RF, Berridge MJ, Schulz I. Release of Ca2+ from a nonmitochondrial intracellular store in pancreatic acinar cells by inositol-1,4,5-trisphosphate. Nature. 1983;306(5938):67-69. — doi:10.1038/306067a0 · PMID: 6605482
- Berridge MJ, Irvine RF. Inositol trisphosphate, a novel second messenger in cellular signal transduction. Nature. 1984;312(5992):315-321. — doi:10.1038/312315a0
- Croze ML, Soulage CO. Potential role and therapeutic interests of myo-inositol in metabolic diseases. Biochimie. 2013;95(10):1811-1827. — doi:10.1016/j.biochi.2013.05.011 · PMID: 23764390
- Bizzarri M, Carlomagno G. Inositol: history of an effective therapy for Polycystic Ovary Syndrome. European Review for Medical and Pharmacological Sciences. 2014;18(13):1896-1903. — PMID: 25010620
- Inositol — history, isolation, and nomenclature — PubMed: inositol history and isolation
- Phosphoinositide signalling and the inositol second-messenger system — PubMed: phosphoinositide and IP3 signalling
External Authoritative Resources
- Sir Michael J. Berridge — biography, IP3 discovery, and Lasker / Wolf / Shaw prizes
- PubChem — myo-Inositol compound record
- PubMed — All research on inositol
Connections
- Inositol
- Inositol Benefits
- All Vitamins
- Choline
- Vitamin B7 (Biotin)
- Vitamin B1 (Thiamine)
- PCOS
- Selenium