Leucine for Muscle Protein Synthesis
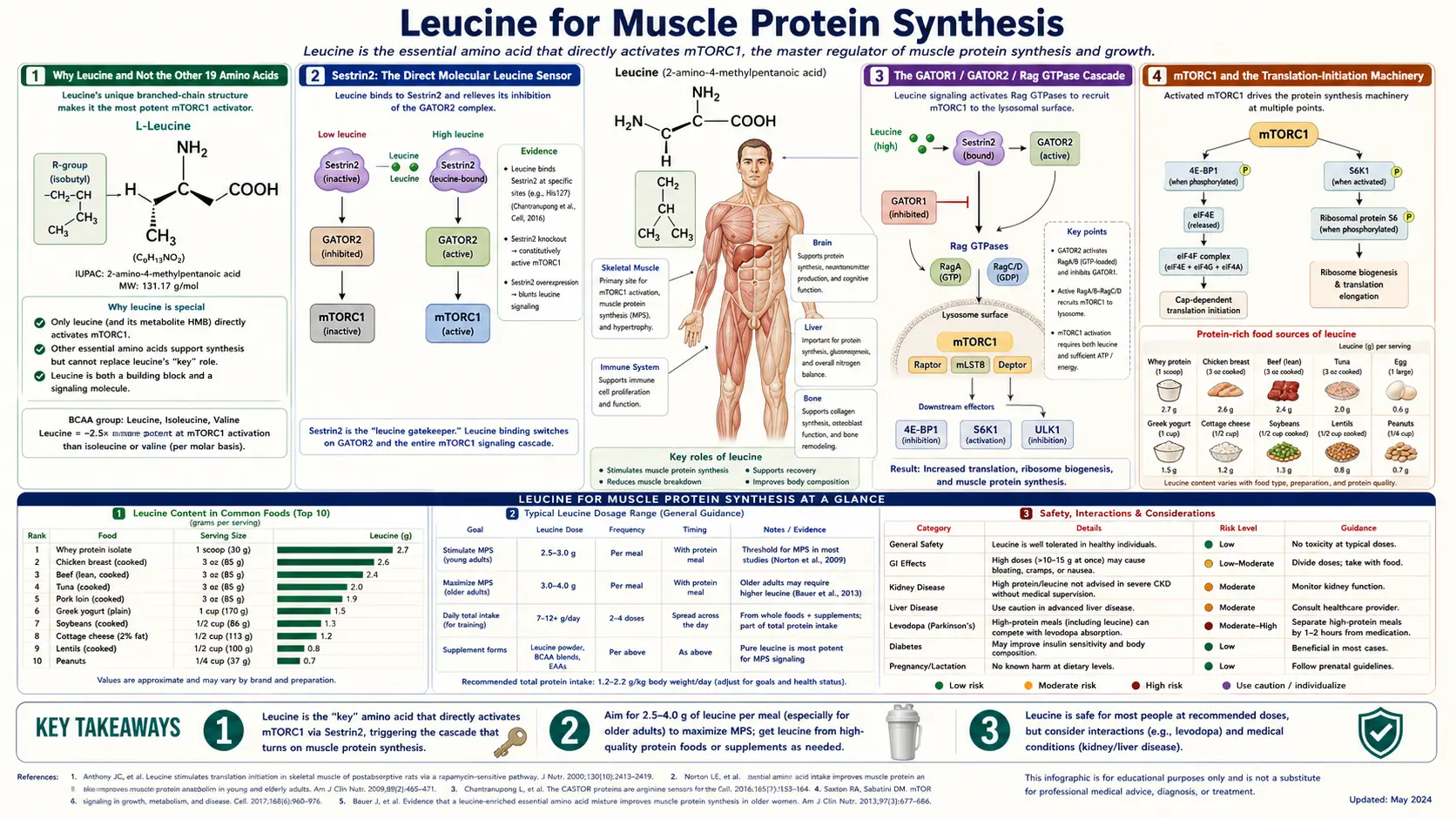
Of the twenty proteinogenic amino acids, leucine is the single most potent nutrient trigger for muscle protein synthesis (MPS). The mechanism — mapped to atomic resolution in the past decade — runs through the lysosomal protein Sestrin2, which directly binds free leucine and releases its inhibition on the GATOR2 complex, allowing the Rag GTPases to recruit mTORC1 to the lysosomal surface where it phosphorylates 4E-BP1, S6K1, and other translation-initiation factors. A practical consequence: a single meal must deliver roughly 2.5–3 g of leucine to fully saturate the MPS response, a threshold met by 25–30 g of whey protein, 30–35 g of beef, or about 35–40 g of egg white — and missed by most cereal-based or low-protein vegetarian meals. This deep-dive walks through the Sestrin2/GATOR molecular sensing mechanism (Wolfson 2016 Nature), the leucine threshold concept developed by Volpi, Wolfe, and Paddon-Jones, why whey concentrate dominates plant proteins for MPS, and the clinical applications across young athletes, post-surgical patients, and the anabolic-resistant elderly.
Table of Contents
- Why Leucine and Not the Other 19 Amino Acids
- Sestrin2: The Direct Molecular Leucine Sensor
- The GATOR1 / GATOR2 / Rag GTPase Cascade
- mTORC1 and the Translation-Initiation Machinery
- The 2.5–3 g Leucine Threshold
- Whey vs Casein vs Plant Proteins
- The Muscle-Full Effect and Refractory Period
- Per-Meal Protein Distribution Across the Day
- The Volpi and Wolfe Research Program
- Clinical Applications and Edge Cases
- Key Research Papers
- Connections
- Featured Videos
Why Leucine and Not the Other 19 Amino Acids
Muscle protein synthesis requires the full complement of all twenty proteinogenic amino acids as building blocks. If any one of the nine essential amino acids is in short supply, ribosomes stall during translation and the new protein is not assembled. Yet only one amino acid — leucine — functions as a signaling molecule that switches on the protein-synthesis machinery itself. Adding leucine to a mixed amino acid infusion produces a measurable spike in MPS; adding equivalent doses of other essential amino acids (isoleucine, valine, lysine, threonine) does not. This signaling specificity makes leucine unique among the twenty.
The selectivity has a deep evolutionary rationale. Leucine is the most abundant amino acid in mammalian muscle protein (roughly 8–10% of total amino acid mass), so a rise in free plasma leucine is a reliable indicator that dietary protein has been consumed in adequate quantity. The cell uses leucine concentration as a proxy for "enough total protein has arrived to make synthesizing new muscle worthwhile" — an elegant solution to the problem of sensing aggregate protein status from a single molecular signal.
The discovery that leucine specifically (not branched-chain amino acids generally, not essential amino acids collectively) carries the signal traces to the 1970s observations of Buse and Reid that leucine infusion stimulated protein synthesis in perfused skeletal muscle. The molecular mechanism remained obscure for forty years until David Sabatini's lab at the Whitehead Institute identified Sestrin2 as the direct leucine-binding sensor in 2016 (Wolfson et al., Nature).
Sestrin2: The Direct Molecular Leucine Sensor
Sestrin2 is a cytosolic protein of approximately 480 amino acids with a deep hydrophobic pocket that binds a single molecule of free leucine with high specificity. The crystal structure (Saxton et al., Science 2016) revealed that leucine sits buried in the pocket making contacts with residues Thr374, Thr377, Glu451, and Trp444 — mutations at these residues abolish leucine binding while preserving the protein fold, allowing direct genetic testing of the sensing hypothesis.
The binding affinity (Kd roughly 20 µM) sits precisely in the physiologically relevant range. Resting plasma leucine concentration in healthy adults is approximately 100–200 µM; postprandial leucine after a protein-rich meal can rise to 400–600 µM. At resting concentrations, only a fraction of Sestrin2 is occupied; at postprandial concentrations, occupancy approaches saturation. This dynamic range gives the sensor its switch-like behavior — below the threshold MPS is constitutive and low; above the threshold MPS is maximally stimulated.
Sestrin2's downstream output is its constitutive inhibition of the GATOR2 complex, a five-subunit pentamer (WDR24, WDR59, MIOS, SEH1L, SEC13). When leucine is absent, Sestrin2 binds GATOR2 and blocks its function. When leucine binds Sestrin2, the protein undergoes a conformational change that releases GATOR2, freeing GATOR2 to inhibit GATOR1, the upstream inhibitor of the Rag GTPases. The double-negative cascade (leucine inhibits Sestrin2 inhibition of GATOR2 which inhibits GATOR1 inhibition of Rags which recruit mTORC1) is a textbook example of regulated relief from constitutive suppression.
The GATOR1 / GATOR2 / Rag GTPase Cascade
GATOR1 is a three-subunit complex (DEPDC5, NPRL2, NPRL3) that functions as a GTPase-activating protein (GAP) for the Rag GTPase heterodimer. The Rag GTPases (RagA or RagB paired with RagC or RagD) cycle between an active and inactive nucleotide-bound state. When GATOR1 is active, it accelerates GTP hydrolysis on RagA/B, locking the Rags into the inactive state. In this state, mTORC1 cannot be recruited to the lysosomal surface and remains diffuse in the cytoplasm, unable to phosphorylate its substrates.
GATOR2 is the upstream brake on GATOR1. When GATOR2 is freed from Sestrin2 inhibition by leucine binding, GATOR2 in turn inhibits GATOR1, allowing the Rags to load GTP and assume the active conformation. Active RagA/B then recruits mTORC1 to the lysosome via direct binding to the Raptor subunit of mTORC1.
At the lysosomal surface, mTORC1 encounters its activator Rheb (Ras homolog enriched in brain), itself regulated by the upstream TSC1/TSC2 complex which integrates growth-factor signaling (insulin, IGF-1) through the PI3K/AKT pathway. The convergence of amino-acid signaling (leucine through Sestrin2/GATOR/Rag) and growth-factor signaling (insulin/IGF-1 through PI3K/AKT/TSC) at the lysosome ensures that mTORC1 fires only when both nutrient and hormone signals are favorable — a built-in safety against runaway anabolism in nutrient-poor conditions.
mTORC1 and the Translation-Initiation Machinery
Active mTORC1 at the lysosomal surface phosphorylates two principal substrates that together control the rate of mRNA translation: 4E-BP1 (eukaryotic translation initiation factor 4E-binding protein 1) and S6K1 (ribosomal protein S6 kinase 1).
4E-BP1 in its dephosphorylated state binds and sequesters eIF4E, the cap-binding protein that recruits ribosomes to the 5′ end of mRNAs. mTORC1-mediated phosphorylation of 4E-BP1 (at Thr37/46, Ser65, Thr70) causes 4E-BP1 to release eIF4E, freeing eIF4E to assemble the eIF4F initiation complex (eIF4E + eIF4G + eIF4A) on capped mRNAs. This step is rate-limiting for cap-dependent translation, particularly for mRNAs with structured 5′ UTRs — which preferentially encode growth-related proteins.
S6K1 phosphorylation activates the kinase, which then phosphorylates ribosomal protein S6 and eIF4B, increasing ribosomal recruitment of mRNAs and enhancing the helicase activity of eIF4A. S6K1 also feeds back to phosphorylate IRS-1, providing a negative-feedback loop to insulin signaling — a mechanism implicated in nutrient-driven insulin resistance.
The net effect is that leucine availability, sensed by Sestrin2 and transduced through GATOR/Rag/mTORC1, increases the rate of cap-dependent translation by roughly 2- to 4-fold within 30–60 minutes. Muscle protein synthesis rates measured by stable-isotope tracer methods (typically L-[ring-13C6]phenylalanine) rise from a fasting baseline of roughly 0.04%/h to a postprandial peak of 0.08–0.10%/h.
The 2.5–3 g Leucine Threshold
The "leucine threshold" hypothesis emerged from a series of careful dose-response studies in healthy young adults and elderly subjects, most prominently by Elena Volpi and Robert Wolfe at the University of Texas Medical Branch and Galveston, and by Stuart Phillips at McMaster University. The core finding: muscle protein synthesis follows a step function, not a continuous dose-response, when plotted against per-meal leucine content. Below approximately 1.7 g of leucine per meal, MPS rises minimally above fasting. Between 1.7 and 2.5 g, MPS rises sharply. Above 2.5 g, MPS plateaus — additional leucine does not produce additional MPS within a single meal.
For young adults, the threshold sits near 1.7–2.0 g of leucine per meal, corresponding to roughly 20–25 g of high-quality protein. For older adults with anabolic resistance, the threshold rises to approximately 2.5–3.0 g of leucine per meal, corresponding to roughly 30–40 g of high-quality protein — a substantially higher per-meal requirement than dietary guidelines typically reflect.
The threshold concept reframes protein recommendations: rather than asking "how many total grams of protein per day," the more clinically meaningful question is "how many meals reach the leucine threshold each day." A patient consuming 60 g of total protein distributed as 10 g at breakfast, 20 g at lunch, and 30 g at dinner achieves the threshold only once (at dinner), generating one MPS event per day. The same 60 g distributed as 20 g per meal achieves the threshold three times, generating three MPS events per day — a meaningful difference in cumulative anabolic response.
Practical translation: high-quality protein sources delivering 2.5–3 g of leucine per serving include 25 g of whey protein isolate, 30 g of beef or chicken, 4 large eggs, 350 g of Greek yogurt, or 200 g of canned tuna. Plant-only sources reach the threshold only with larger portions (roughly 50 g of soy protein isolate, 150 g of cooked lentils plus 100 g of cooked quinoa).
Whey vs Casein vs Plant Proteins
Not all proteins are equivalent in their capacity to stimulate MPS, even when matched for total protein content. Three properties matter: leucine concentration (percentage of leucine in the protein), digestion kinetics (how quickly amino acids appear in circulation), and protein quality (the presence of all essential amino acids in physiologically useful ratios).
Whey protein isolate is the gold standard. Leucine content is approximately 10–11% of total protein (the highest of any common food protein). Digestion is rapid — whey is liquid at gastric pH, empties from the stomach within 15–30 minutes, and produces peak plasma leucine within 60–90 minutes of ingestion. A 25 g serving of whey isolate delivers approximately 2.5–3 g of leucine and reliably exceeds the MPS threshold in young adults.
Casein contains a similar leucine percentage (approximately 9–10%) but precipitates in gastric acid, forming a clot that is digested over 4–6 hours. The slow appearance of amino acids produces a lower peak plasma leucine (insufficient to saturate the threshold) but sustained delivery over hours. Casein is therefore preferred for overnight protein dosing and for situations where prolonged amino acid availability matters more than peak MPS stimulation.
Egg white protein sits between whey and casein on the leucine scale (approximately 8.5% leucine) with moderate digestion kinetics. A 30 g serving delivers approximately 2.5 g of leucine.
Soy protein isolate is the highest-leucine plant protein (approximately 8% leucine) and the only plant protein routinely capable of reaching the MPS threshold at standard serving sizes. A 30 g serving delivers approximately 2.4 g of leucine — marginally below the young-adult threshold and meaningfully below the elderly threshold. Soy isolate is the practical first choice for plant-based athletes.
Other plant proteins (pea, rice, hemp) have leucine percentages in the 7–8% range but suffer from low total protein density — reaching 2.5 g of leucine requires 35–40 g of isolate or much larger food portions. Pea protein with added free leucine (typically 2–3 g per serving) is one solution that has been tested in clinical trials.
The practical recommendation for plant-only diets: combine pea or soy isolate with a small bolus of free leucine (1–2 g) to reliably exceed the threshold, particularly in older adults.
The Muscle-Full Effect and Refractory Period
A counterintuitive finding from continuous-infusion stable-isotope studies: continuously elevated plasma leucine does not produce continuously elevated MPS. After approximately 90–120 minutes of leucine-driven MPS, the muscle becomes refractory to further anabolic stimulation — the "muscle-full effect" described by Atherton and Rennie at the University of Nottingham.
Mechanistically, the muscle-full effect involves at least three layers of negative feedback: S6K1-mediated phosphorylation of IRS-1 that blunts insulin signaling; mTORC1-mediated phosphorylation of GRB10 that further attenuates growth-factor signaling; and accumulation of unfolded protein in the endoplasmic reticulum triggering the unfolded protein response and translational arrest.
The practical consequence is that increasing per-meal leucine beyond the threshold does not increase the magnitude or duration of the MPS response. Spacing protein-rich meals by 3–4 hours allows the muscle to exit the refractory state and respond to the next leucine bolus. Eating six small protein meals spaced 2 hours apart is therefore inferior to eating three protein-rich meals spaced 4–5 hours apart — the latter generates three full MPS events; the former generates one full event plus several blunted ones.
Per-Meal Protein Distribution Across the Day
Most Americans consume their protein in a skewed distribution: low at breakfast (often 5–10 g, dominated by carbohydrates), moderate at lunch (15–25 g), and concentrated at dinner (35–50 g). This pattern reaches the leucine threshold once per day (at dinner), generating one MPS event.
The "even-distribution hypothesis" tested by Mamerow and colleagues (Journal of Nutrition 2014) compared 90 g/day of protein distributed as 30 g per meal vs the typical skewed distribution. The even-distribution arm produced approximately 25% higher 24-hour MPS, validating the per-meal threshold model. Subsequent work by Murphy, Phillips, and others has confirmed and extended these findings in athletic populations and the elderly.
For practical implementation: target 25–30 g of high-quality protein at breakfast, lunch, and dinner. Eggs, Greek yogurt, cottage cheese, smoked salmon, or a whey-based smoothie are practical breakfast options that reach the leucine threshold. Skipping breakfast or relying on toast-and-coffee patterns is one of the most easily corrected sources of suboptimal protein status in otherwise healthy adults.
The Volpi and Wolfe Research Program
Elena Volpi and Robert Wolfe's laboratories (initially at the University of Texas Medical Branch, then at the University of Arkansas) have generated the most influential body of human stable-isotope tracer work on leucine and MPS over the past two decades. Their core methodology infuses a labeled amino acid (typically L-[ring-13C6]phenylalanine or L-[13C1]leucine) to steady state, samples plasma and muscle biopsies, and calculates muscle protein fractional synthetic rate from the precursor-product ratio.
Key findings from this program: (1) Free leucine bolus alone (without other amino acids) stimulates MPS for approximately 30 minutes before plateauing — the response requires the full essential amino acid complement to sustain. (2) Anabolic resistance in the elderly is overcome by raising per-meal leucine to 2.5–3 g, restoring MPS to young-adult levels. (3) Co-ingestion of carbohydrate with protein does not enhance MPS beyond protein alone in adequately-protein-fed subjects — the insulin spike is permissive but not synergistic. (4) Resistance exercise sensitizes muscle to leucine for 24–48 hours post-exercise, lowering the threshold and amplifying the response.
These findings have been incorporated into the PROT-AGE consensus recommendations (Bauer et al. 2013, JAMDA) that older adults should consume 1.0–1.2 g protein per kg body weight per day, distributed as at least 25–30 g per meal — substantially higher than the legacy RDA of 0.8 g/kg/day that was derived from nitrogen-balance studies in young subjects.
Clinical Applications and Edge Cases
- Bariatric surgery patients — restrictive procedures (sleeve gastrectomy, Roux-en-Y gastric bypass) limit meal volume below what is needed to reach the leucine threshold from whole food. Whey or essential amino acid supplementation is standard to preserve lean mass during rapid weight loss.
- Intensive care patients — critical illness drives accelerated muscle catabolism (ICU-acquired weakness affects 50–80% of long-stay patients). High-leucine enteral formulas and parenteral amino acid solutions enriched in leucine are an active area of trial.
- Cancer cachexia — advanced cancer produces a syndrome of accelerated muscle loss driven by tumor-derived cytokines and metabolic dysregulation. Leucine and HMB have been tested as anti-catabolic adjuncts with modest but reproducible benefit on lean mass preservation.
- Astronauts and bed-rest models — microgravity and immobilization produce profound anabolic resistance. Leucine-enriched essential amino acid supplementation is part of NASA countermeasure protocols.
- Maple syrup urine disease (MSUD) — the rare inborn error of branched-chain alpha-keto acid dehydrogenase. Patients cannot metabolize leucine; supplementation is contraindicated and dietary leucine must be tightly restricted to prevent toxic accumulation and neurologic injury.
- Chronic kidney disease (advanced) — protein restriction is sometimes prescribed in pre-dialysis CKD to slow progression. The trade-off against accelerated sarcopenia is genuine; ketoacid analogs of leucine that bypass the nitrogen burden have been tested as a compromise.
For most healthy adults, the practical implementation is simpler than the underlying biochemistry suggests: eat 25–30 g of high-quality protein at each of three meals, train resistance 2–4 times per week, and the system delivers maximal MPS without supplementation. The need for free-form leucine arises mainly when whole-food protein cannot reach the threshold (elderly with low appetite, bariatric surgery, strict vegan diets, or critical illness).
Key Research Papers
- Wolfson RL et al. (2016). Sestrin2 is a leucine sensor for the mTORC1 pathway. Nature. — PubMed
- Saxton RA et al. (2016). Structural basis for leucine sensing by the Sestrin2-mTORC1 pathway. Science. — PubMed
- Anthony JC et al. (2000). Leucine stimulates translation initiation in skeletal muscle of postabsorptive rats via a rapamycin-sensitive pathway. Journal of Nutrition. — PubMed
- Volpi E et al. (2003). Essential amino acids are primarily responsible for the amino acid stimulation of muscle protein anabolism in healthy elderly adults. American Journal of Clinical Nutrition. — PubMed
- Atherton PJ, Rennie MJ (2010). It's no longer just about "the muscle full effect". Journal of Physiology. — PubMed
- Mamerow MM et al. (2014). Dietary protein distribution positively influences 24-h muscle protein synthesis in healthy adults. Journal of Nutrition. — PubMed
- Moore DR et al. (2009). Ingested protein dose response of muscle and albumin protein synthesis after resistance exercise in young men. American Journal of Clinical Nutrition. — PubMed
- Tang JE et al. (2009). Ingestion of whey hydrolysate, casein, or soy protein isolate: effects on mixed muscle protein synthesis at rest and following resistance exercise. Journal of Applied Physiology. — PubMed
- Boirie Y et al. (1997). Slow and fast dietary proteins differently modulate postprandial protein accretion. PNAS. — PubMed
- Norton LE et al. (2009). The leucine content of a complete meal directs peak activation but not duration of skeletal muscle protein synthesis. Journal of Nutrition. — PubMed
- Bauer J et al. (2013). Evidence-based recommendations for optimal dietary protein intake in older people: PROT-AGE Study Group. JAMDA. — PubMed
- Phillips SM (2014). A brief review of critical processes in exercise-induced muscular hypertrophy. Sports Medicine. — PubMed
PubMed Topic Searches
- PubMed: Leucine MPS and mTORC1
- PubMed: Sestrin2 / GATOR2 leucine sensing
- PubMed: Leucine threshold per meal
- PubMed: Whey vs casein vs soy MPS
- PubMed: Per-meal protein distribution
Connections
- Leucine Benefits Hub
- Leucine Overview
- mTOR Activation
- Sarcopenia Prevention
- Recovery and Exercise
- Isoleucine
- Valine
- Glutamine
- Rapamycin (mTOR Inhibitor)
- Creatine
- Collagen
- Eggs (Whey, Casein, Albumin)
- Beef
- Insulin Resistance
- All Amino Acids