Sirtuin Activators for Longevity
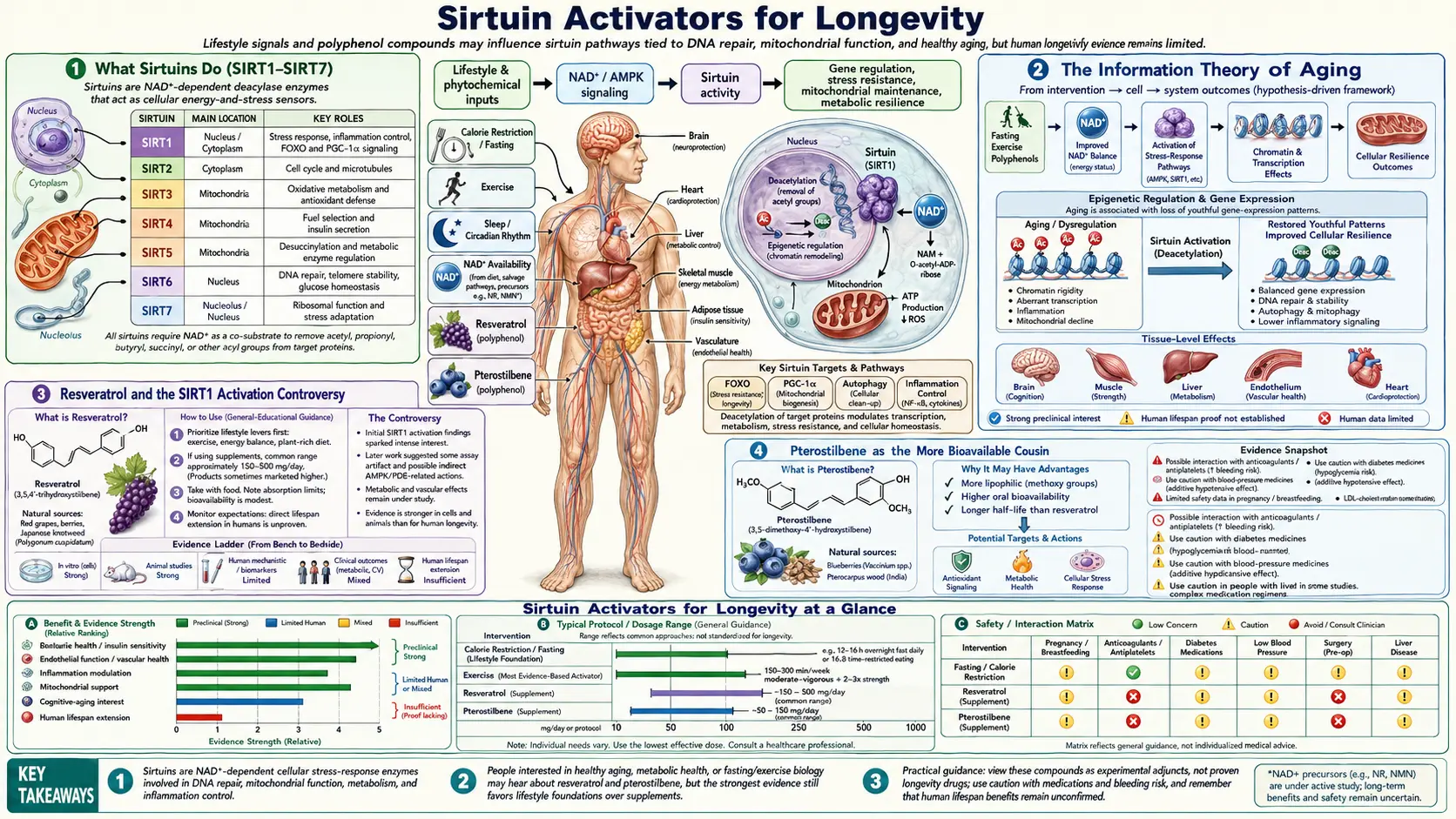
Sirtuins are a family of seven mammalian NAD+-dependent deacylases (SIRT1–SIRT7) that act as master regulators of metabolism, DNA repair, and the response to nutrient stress. They were identified as longevity genes in yeast in 2000 by Leonard Guarente's lab at MIT, and the discovery that resveratrol activated yeast SIRT2 (Sir2) ignited a fifteen-year wave of pharmaceutical and consumer interest in "STACs" (sirtuin-activating compounds). The story has since become more complicated — a major 2010 paper argued that the original resveratrol-SIRT1 activation was a fluorophore artifact, and the GSK acquisition of Sirtris for $720M in 2008 was followed by the eventual shuttering of its STAC pipeline. The current evidence is more nuanced than either the boosters or the skeptics suggest: sirtuins are real longevity regulators, NAD+-driven sirtuin activation is well-supported, but small-molecule allosteric activators like resveratrol and pterostilbene act on a more limited subset of substrates than originally claimed.
Table of Contents
- What Sirtuins Do (SIRT1–SIRT7)
- The Information Theory of Aging
- Resveratrol and the SIRT1 Activation Controversy
- Pterostilbene as the More Bioavailable Cousin
- Second-Generation STACs (SRT2104, SRT1720)
- Sirtuin Activation Through Substrate (NAD+)
- SIRT6 Overexpression Extends Mouse Lifespan
- Human Evidence for Resveratrol
- Practical Application and Cautions
- Key Research Papers
- Connections
- Featured Videos
What Sirtuins Do (SIRT1–SIRT7)
The mammalian sirtuin family has seven members with overlapping but distinct functions. They share the same catalytic core: removing acyl groups (most commonly acetyl, but also succinyl, malonyl, and others) from lysine residues on target proteins, using NAD+ as an obligate cosubstrate. Each deacylation reaction consumes one NAD+ molecule, which is why sirtuin activity is fundamentally constrained by NAD+ availability.
- SIRT1 (nuclear): the most studied sirtuin. Deacetylates p53 (regulating apoptosis), FOXO transcription factors (regulating stress response), PGC-1alpha (regulating mitochondrial biogenesis), NF-kB (regulating inflammation), and many others. Acts as a central node in the response to caloric restriction.
- SIRT2 (cytoplasmic, nuclear): regulates microtubule dynamics, cell cycle, and adipogenesis.
- SIRT3 (mitochondrial): the major mitochondrial deacetylase. Activates the antioxidant enzyme MnSOD (SOD2) and many enzymes of the TCA cycle and beta-oxidation. SIRT3 activity declines with age and is implicated in age-related metabolic dysfunction.
- SIRT4 (mitochondrial): ADP-ribosylates glutamate dehydrogenase, regulating amino acid metabolism.
- SIRT5 (mitochondrial): removes succinyl, malonyl, and glutaryl groups (not acetyl) from a broad range of mitochondrial enzymes.
- SIRT6 (nuclear, chromatin): regulates DNA double-strand-break repair, telomere maintenance, and chromatin stability. SIRT6 overexpression dramatically extends male mouse lifespan (Kanfi et al., Nature 2012).
- SIRT7 (nucleolar): regulates ribosomal RNA transcription and nucleolar function.
The seven sirtuins together affect essentially every cellular process implicated in aging — energy metabolism, DNA repair, mitochondrial function, oxidative stress response, inflammation, and chromatin maintenance. This breadth is what makes them attractive longevity targets and what makes "sirtuin activation" a marketing-friendly but technically vague claim.
The Information Theory of Aging
David Sinclair's 2014 book "Lifespan" and the underlying scientific papers articulate a unifying framework called the "Information Theory of Aging." The theory says that aging is primarily a loss of epigenetic information — the chromatin marks (DNA methylation, histone acetylation, histone methylation) that tell each cell which subset of its genome to express.
The Sinclair lab's ICE (Induced Changes to Epigenome) mouse experiments, published in Cell in 2023, showed that repeated induction of DNA double-strand breaks (without changing the DNA sequence itself) produced an accelerated-aging phenotype in mice — reduced lifespan, gray hair, frailty, all the standard markers of aging. Because the DNA sequence was unchanged, the mechanism was attributed to chromatin remodeling and epigenetic drift — the cells "forgot" which cell type they were.
Within this framework, sirtuins are essential because they are recruited to sites of DNA damage to facilitate repair, and the repeated relocation away from their normal chromatin sites is what drives the epigenetic drift. NAD+ availability constrains sirtuin recruitment. Therefore, restoring NAD+ (with NMN or NR) and activating sirtuins (with resveratrol or pterostilbene) is theorized to slow the loss of epigenetic information.
The Information Theory is not universally accepted. Critics point out that DNA methylation clocks (Horvath, GrimAge, DunedinPACE) measure epigenetic patterns but do not prove causation — an epigenetic clock that ticks faster in stressed cells does not mean that resetting the clock would restore youth. The Sinclair lab's subsequent partial-reprogramming work (using three of the four Yamanaka factors, OSK, to reset epigenetic age in mouse retinal cells) is provocative but early. The framework is influential, but the clinical translation is mostly speculative.
Resveratrol and the SIRT1 Activation Controversy
Resveratrol is a stilbenoid polyphenol found in grape skins, red wine, and Japanese knotweed (the commercial source). The Sirtris story begins with Howitz et al. in Nature 2003, where the Sinclair lab reported that resveratrol activated yeast Sir2 (SIRT1 homolog) and extended yeast replicative lifespan. Subsequent work in worms and flies showed similar effects, and the David Sinclair group's 2006 Nature paper (Baur et al.) reported that resveratrol improved survival and metabolic health of mice on a high-calorie diet — one of the most cited animal longevity papers of the 2000s.
The activation mechanism was reported as direct allosteric activation of SIRT1 — resveratrol binding the enzyme and increasing its substrate affinity. This finding led GSK to acquire Sirtris in 2008 for $720M and to develop more potent synthetic STACs.
The trouble started in 2010 with Pacholec et al. (J Biol Chem), who showed that resveratrol's apparent SIRT1 activation in vitro was an artifact of the fluorophore-tagged peptide substrate used in standard assays. When tested against unmodified protein substrates, resveratrol showed little or no SIRT1 activation. The Pacholec paper concluded that resveratrol and the Sirtris-developed STACs (SRT2183, SRT1460, SRT1720) were not direct SIRT1 activators.
The Sinclair group's 2013 Science paper (Hubbard et al.) responded that resveratrol DOES activate SIRT1 directly, but only on a specific subset of substrates whose target lysines are flanked by hydrophobic residues. The mechanism is "assisted allosteric activation" rather than universal activation. Hubbard et al. identified PGC-1alpha and FOXO3 as substrates whose deacetylation IS accelerated by resveratrol in vivo. So both the original claim (resveratrol activates SIRT1) and the artifact critique (the original assay was misleading) turn out to be partially correct: resveratrol is a substrate-selective SIRT1 activator.
The clinical implication is that resveratrol's sirtuin-activating effect is real but more limited than the marketing implies. It may activate SIRT1 toward a small number of important substrates without globally turning on all SIRT1 activity. Whether this is clinically meaningful at the gram doses used in human trials remains uncertain.
Pterostilbene as the More Bioavailable Cousin
Pterostilbene is a dimethylated analog of resveratrol found in blueberries and Indian kino tree. The two structural changes — replacing two of the three hydroxyl groups with methyl ethers — dramatically alter pharmacokinetics. Pterostilbene has approximately 4× longer plasma half-life, 8× higher oral bioavailability, and crosses the blood-brain barrier more readily than resveratrol.
For these reasons, pterostilbene is the preferred stilbenoid in many longevity formulations. The Niagen+ formula from ChromaDex combines NR with pterostilbene specifically. Anti-aging effects in mice have been demonstrated for pterostilbene (Mitchell et al., Cell Reports 2014, showed pterostilbene improved healthspan in male mice on a Western diet).
Human trial data are limited. A 2016 trial (Riche et al., J Toxicol) found that 250 mg/day pterostilbene reduced LDL cholesterol and blood pressure modestly over 6–8 weeks. No long-term endpoint data exist. Side effects appear minimal.
The honest comparison: pterostilbene is probably what people who supplement resveratrol should actually be taking, given the superior pharmacokinetics. The cost is roughly 2–3× that of resveratrol per gram, and few products contain it at meaningful doses.
Second-Generation STACs (SRT2104, SRT1720)
After the Sirtris/GSK acquisition, the pharmaceutical pipeline focused on synthetic STACs — small molecules with greater potency and selectivity than resveratrol. The leading compounds were SRT1720, SRT2104, and SRT2183.
SRT1720 showed striking results in obese mice (Minor et al., Sci Rep 2011): improved insulin sensitivity, increased mitochondrial biogenesis, extended lifespan on high-fat diet. SRT2104 entered clinical trials and was studied in metabolic disease, psoriasis, and ulcerative colitis. The results were disappointing — modest or no improvement in most endpoints, and the synthetic STAC program was largely shelved.
The reasons for the clinical disappointment likely include: human SIRT1 activity may not be the rate-limiting factor for the clinical outcomes targeted; the chronic safety profile of long-term sirtuin activation in humans is unknown; and the trials used short-term endpoints in patients with established disease, which may not be where sirtuin activation would help.
The synthetic STAC story is a cautionary tale about extrapolating from rodent longevity assays to human clinical endpoints. The mechanism worked as advertised in mice; the clinical translation did not.
Sirtuin Activation Through Substrate (NAD+)
The most reliable way to increase sirtuin activity is not through allosteric activators but by raising NAD+ availability. Sirtuin activity is rate-limited by intracellular NAD+ in most conditions, especially in older tissues where NAD+ has declined. Raising NAD+ by 50% should therefore raise sirtuin activity proportionally, regardless of whether an allosteric activator is also present.
This is why the NAD Boosters page and this page are complementary — NAD precursors provide the substrate, and stilbenoids (if they work) increase the catalytic efficiency on selected substrates. The combination, in theory, should be more effective than either alone.
Caloric restriction acts through both mechanisms simultaneously: it reduces glycolytic NADH production (raising the NAD+/NADH ratio) and reduces nutrient signals that suppress sirtuin activity. This is one of the reasons CR is the strongest sirtuin activator known. The pharmaceutical longevity strategy of combining NR/NMN with resveratrol/pterostilbene is essentially trying to reproduce the CR signal without the food restriction.
SIRT6 Overexpression Extends Mouse Lifespan
The most striking sirtuin longevity finding outside of SIRT1 is the SIRT6 overexpression work from the Cohen lab at Bar-Ilan (Kanfi et al., Nature 2012). Transgenic male mice overexpressing SIRT6 (but not female mice) had approximately 15% extended median lifespan compared to wild-type littermates. The extension was attributed to reduced IGF-1 signaling and improved metabolic homeostasis.
SIRT6 has multiple essential functions: it deacetylates histone H3K9ac and H3K56ac at telomeres and DNA repair sites, it ADP-ribosylates PARP1 to activate DNA double-strand-break repair, and it represses glycolysis at the transcriptional level. SIRT6 deficiency in mice causes a severe progeroid phenotype with death by 4 weeks.
Small molecule SIRT6 activators are an active area of drug discovery. UBCS039 and MDL-800 are early-stage compounds with reasonable in vitro potency. None have entered clinical trials. Anthocyanins (the pigments in blueberries and dark berries) have been reported to activate SIRT6 in vitro, but the in vivo evidence is weak.
The SIRT6 story is one of the strongest pieces of evidence that sirtuin pathway manipulation can extend mammalian lifespan when done at the right point in the pathway with the right tools.
Human Evidence for Resveratrol
Resveratrol has been extensively studied in human trials — far more than any other STAC. The literature is large enough to draw some conclusions.
- Cardiovascular endpoints: meta-analyses of resveratrol trials (Liu et al., Am J Clin Nutr 2015) show modest reductions in systolic blood pressure (~3–5 mmHg) and improvements in flow-mediated dilation. Effects are small but reasonably consistent.
- Diabetes: meta-analyses show modest reductions in fasting glucose and HbA1c in type 2 diabetics on resveratrol. Effect size is small (~0.2% HbA1c reduction).
- Inflammation: variable effects on CRP and TNF-alpha, generally in the direction of reduction but inconsistently.
- Aging biomarkers: no published trial has demonstrated epigenetic clock improvement on resveratrol.
- Lifespan: no human trial has reported survival data.
- Notable null results: the InCHIANTI cohort study (Semba et al., JAMA Intern Med 2014) tracked urinary resveratrol metabolites in 783 community-dwelling Italian older adults and found NO association between resveratrol exposure (from food and supplements) and cardiovascular events, cancer, or mortality over 9 years. This is one of the strongest pieces of evidence that dietary-level resveratrol intake (the level achievable from red wine, grapes, and supplements at typical doses) does not meaningfully extend life.
The honest summary on resveratrol in humans: modest biomarker effects at 250 mg–1g/day doses, no demonstrated longevity benefit, the InCHIANTI null result strongly suggesting that resveratrol from dietary or supplement sources at achievable doses does not extend life in well-nourished adults. The compound is well-tolerated and inexpensive, but the evidence for it as a longevity intervention is weak.
Practical Application and Cautions
For an individual considering sirtuin-related supplementation:
- Prioritize substrate over allosteric activators: if sirtuin activity matters, raising NAD+ through NR (or NMN, with the caveats on the NAD Boosters page) is more evidence-supported than adding resveratrol on top.
- Pterostilbene over resveratrol: if choosing a stilbenoid, pterostilbene's superior pharmacokinetics and blood-brain-barrier penetration make it the more rational choice at 100–250 mg/day.
- Activate sirtuins through CR and exercise: both raise NAD+ and reduce sirtuin-inhibitory signals. These are the most reliable sirtuin activators.
- Eat the food sources: blueberries (pterostilbene, anthocyanins), red grapes and red wine (resveratrol), Japanese knotweed (concentrated resveratrol), parsley (apigenin, CD38 inhibitor), turmeric (curcumin, sirtuin activator at high doses).
Cautions:
- Resveratrol has estrogenic activity at high doses; women with estrogen-sensitive cancer history should be cautious.
- Resveratrol inhibits CYP3A4 and several other drug-metabolizing enzymes; significant drug interactions are possible at gram doses.
- Resveratrol is a mild blood thinner and may potentiate anticoagulants.
- Most commercial resveratrol products are low-quality due to oxidation during manufacturing and storage; trans-resveratrol is the active form, and many products contain mostly degraded material.
- Sirtuin activation increases mitochondrial biogenesis and DNA repair activity; in cells with established oncogenic mutations, this could theoretically support tumor metabolism. The clinical relevance is unclear.
Key Research Papers
- Tissenbaum HA, Guarente L. Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans. Nature 2001; 410:227–230 — PubMed PMID: 11242085
- Howitz KT et al. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature 2003; 425:191–196 — PubMed PMID: 12939617
- Baur JA et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006; 444:337–342 — PubMed PMID: 17086191
- Pearson KJ et al. Resveratrol delays age-related deterioration and mimics transcriptional aspects of dietary restriction without extending life span. Cell Metab 2008; 8:157–168 — PubMed PMID: 18599363
- Pacholec M et al. SRT1720, SRT2183, SRT1460, and resveratrol are not direct activators of SIRT1. J Biol Chem 2010; 285:8340–8351 — PubMed PMID: 20061378
- Minor RK et al. SRT1720 improves survival and healthspan of obese mice. Sci Rep 2011; 1:70 — PubMed PMID: 22355589
- Kanfi Y et al. The sirtuin SIRT6 regulates lifespan in male mice. Nature 2012; 483:218–221 — PubMed PMID: 22367546
- Hubbard BP et al. Evidence for a common mechanism of SIRT1 regulation by allosteric activators. Science 2013; 339:1216–1219 — PubMed PMID: 23471411
- Mitchell SJ et al. The SIRT3 activator pterostilbene improves healthspan in mice on a Western diet. Cell Rep 2014; 6:836–843 — Search PubMed
- Semba RD et al. Resveratrol levels and all-cause mortality in older community-dwelling adults. JAMA Intern Med 2014; 174:1077–1084 (InCHIANTI null result) — Search PubMed
- Liu Y et al. Effect of resveratrol on blood pressure: a meta-analysis of randomized controlled trials. Clin Nutr 2015; 34:27–34 — PubMed PMID: 24731650
- Imai S, Guarente L. NAD+ and sirtuins in aging and disease. Trends Cell Biol 2014; 24:464–471 — PubMed PMID: 24786309
- Bonkowski MS, Sinclair DA. Slowing ageing by design: the rise of NAD+ and sirtuin-activating compounds. Nat Rev Mol Cell Biol 2016; 17:679–690 — PubMed PMID: 27552971
- Mostoslavsky R et al. Genomic instability and aging-like phenotype in the absence of mammalian SIRT6. Cell 2006; 124:315–329 — PubMed PMID: 16439206
- Yang H et al. Loss of epigenetic information as a cause of mammalian aging. Cell 2023; 186:305–326 (Sinclair ICE mouse) — PubMed PMID: 36638792
Connections
- Benefits Deep Dive (Hub)
- Caloric Restriction and Mimetics
- NAD Boosters
- Sinclair Stack and Critique
- Longevity Protocols (Main)
- Resveratrol
- Pterostilbene
- NMN (Nicotinamide Mononucleotide)
- NR (Nicotinamide Riboside)
- Blueberries
- Japanese Knotweed
- Curcumin
- Apigenin
- Quercetin