Serine as a Glycine Source — SHMT and One-Carbon Metabolism
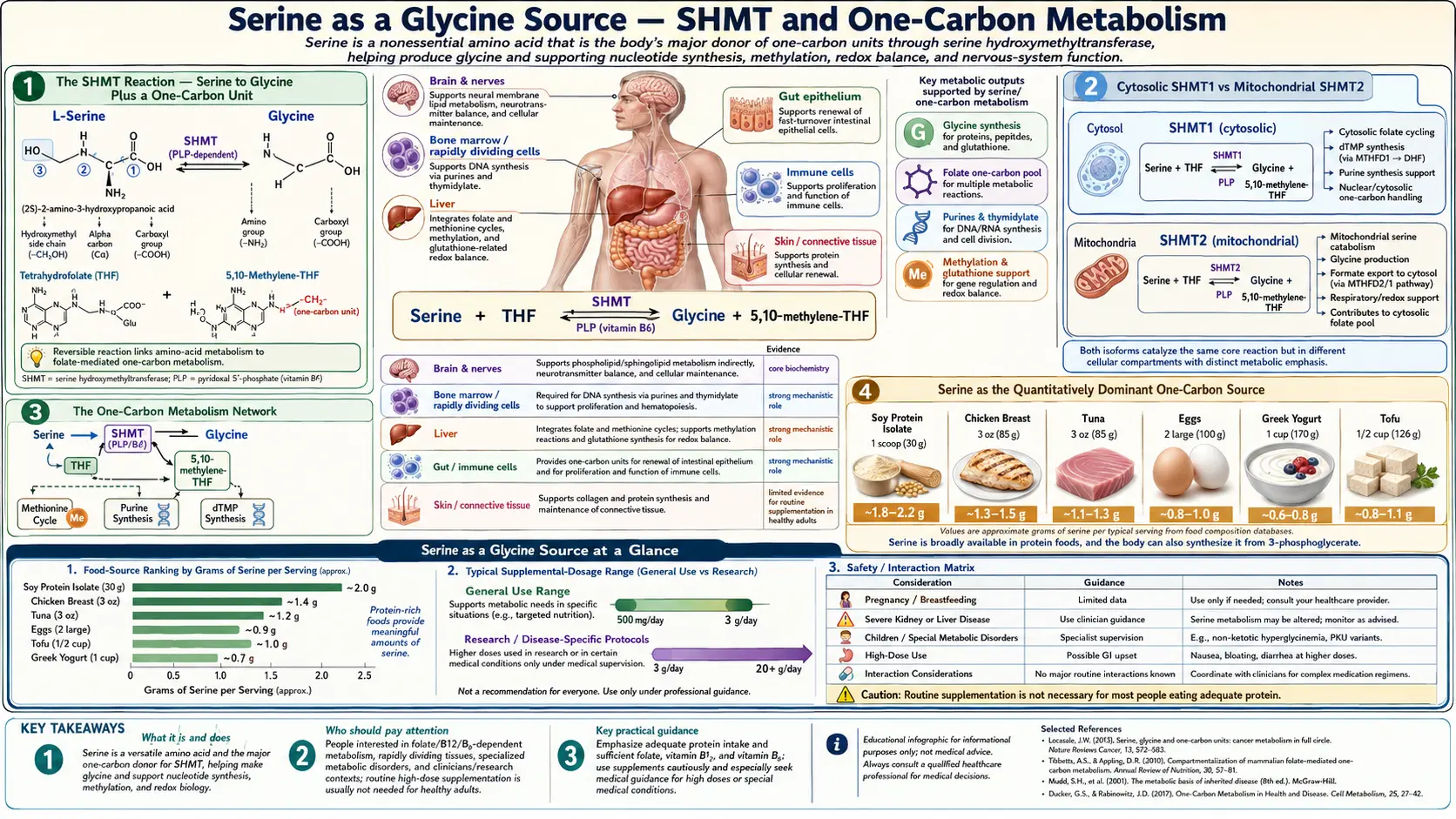
Serine and glycine are interconvertible in a single enzyme reaction catalyzed by serine hydroxymethyltransferase (SHMT). The reaction does two jobs at once: it generates glycine from serine, and it transfers a one-carbon unit from serine to tetrahydrofolate, feeding the entire folate-mediated methylation network. This is the quantitatively dominant source of one-carbon units in human cells, and the rate at which serine flows through SHMT influences DNA methylation, neurotransmitter synthesis, glutathione production, homocysteine clearance, and red-blood-cell production. It also explains a clinical puzzle: even though glycine has an officially recognized RDA, dietary glycine deficiency rarely produces symptoms in healthy adults because serine flux through SHMT compensates. This deep-dive walks through the SHMT reaction, the folate cycle interface, the methylation network, and the clinical implications — from cancer metabolism to MTHFR variants to the dietary glycine question.
Table of Contents
- The SHMT Reaction — Serine to Glycine Plus a One-Carbon Unit
- Cytosolic SHMT1 vs Mitochondrial SHMT2
- The One-Carbon Metabolism Network
- Serine as the Quantitatively Dominant One-Carbon Source
- Interface with the Methionine / SAMe Methylation Cycle
- The Glycine Cleavage System — Glycine as Secondary One-Carbon Source
- Why Dietary Glycine Deficiency Rarely Produces Symptoms
- SHMT and Cancer Metabolism — The Drugging Strategy
- MTHFR Variants and the Methylation-Cycle Genetics
- Clinical Implications — When to Supplement Glycine vs Serine
- Key Research Papers
- Connections
- Featured Videos
The SHMT Reaction — Serine to Glycine Plus a One-Carbon Unit
Serine hydroxymethyltransferase (SHMT, also called serine transhydroxymethylase, EC 2.1.2.1) catalyzes the reversible reaction:
L-serine + tetrahydrofolate (THF) ⇌ glycine + 5,10-methylenetetrahydrofolate (5,10-CH2-THF) + H2O
The reaction does two things simultaneously, and the elegance of the chemistry is worth a moment of attention:
- Cleavage of serine — the hydroxymethyl group at the β-carbon of serine is broken off, converting serine to glycine
- Transfer of the hydroxymethyl carbon to tetrahydrofolate — the released CH2OH unit is captured by THF, producing 5,10-methylene-THF, the activated one-carbon donor that feeds the rest of folate metabolism
The enzyme requires pyridoxal-5'-phosphate (PLP, the active form of vitamin B6) as cofactor. PLP forms a Schiff base with the α-amino group of serine, and the resulting structure stabilizes the carbanion intermediate that breaks the Cα-Cβ bond. Vitamin B6 deficiency therefore reduces SHMT activity in vivo and impairs the serine-to-glycine and serine-to-folate flux, which has secondary consequences for methylation status and DNA synthesis.
The reaction is reversible in principle (glycine plus 5,10-methylene-THF could in theory regenerate serine), and the direction in any given tissue depends on the relative concentrations of the four reactants and on cellular demand. In most cells under most conditions, the reaction runs in the forward direction (serine to glycine) because cellular demand for one-carbon units and glycine exceeds demand for serine, and because parallel pathways consume the products.
The thermodynamics make SHMT one of the central control points of one-carbon metabolism. The standard free energy of the reaction is close to zero, so flux direction is determined by mass action. Cancer cells, which have enormous demand for one-carbon units to support DNA synthesis, drive the reaction strongly forward (consuming serine, producing glycine plus 5,10-methylene-THF). This has therapeutic implications discussed in the cancer metabolism section below.
Cytosolic SHMT1 vs Mitochondrial SHMT2
Mammalian cells express two SHMT isoforms encoded by separate genes:
- SHMT1 (cytosolic, encoded by SHMT1 on chromosome 17p11) — predominant in adult liver, kidney, and most differentiated tissues. Provides cytosolic 5,10-methylene-THF for thymidylate synthesis (DNA precursor) and for the cytosolic arm of the methionine cycle.
- SHMT2 (mitochondrial, encoded by SHMT2 on chromosome 12q13) — expressed in nearly all cell types, particularly in proliferating cells and tumors. Provides mitochondrial 5,10-methylene-THF for the mitochondrial portion of the folate cycle, which is the principal source of formate that is exported to the cytosol to support cytosolic one-carbon metabolism.
Both enzymes catalyze the same chemical reaction but in spatially distinct compartments. The compartmentalization matters because mitochondria and cytosol have separate pools of tetrahydrofolate cofactor (the folate carrier protein SLC25A32 transports specific folate species across the mitochondrial inner membrane). The bulk of cellular one-carbon flux runs through the mitochondrial SHMT2 arm: serine enters mitochondria, is cleaved by SHMT2, and the resulting 5,10-methylene-THF is processed through MTHFD2 and MTHFD1L to generate formate, which is exported to the cytosol.
SHMT2 is highly upregulated in cancer. Many tumors require SHMT2 activity to support their proliferation, and SHMT2 expression correlates with tumor aggressiveness across multiple cancer types. Pharmacological inhibition of SHMT2 is an active area of cancer drug development, with SHIN1 and SHIN2 (small-molecule SHMT inhibitors) showing preclinical activity in lymphoma and other cancers.
The One-Carbon Metabolism Network
"One-carbon metabolism" refers to the network of reactions that transfer single-carbon groups (methyl, methylene, methenyl, formyl) between metabolites, mediated by tetrahydrofolate cofactors. This network produces purine and thymidylate nucleotides for DNA and RNA synthesis, regenerates the methionine pool that feeds the universal methyl donor S-adenosylmethionine (SAMe), and supports the synthesis of glycine, serine, and several other small molecules.
The core players:
- Tetrahydrofolate (THF) — the cofactor backbone, carrying one-carbon units at the N5 and N10 positions of the pteridine ring. Dietary folate (folic acid, leafy greens, fortified flour) enters this pool after reduction by dihydrofolate reductase (DHFR).
- 5,10-methylene-THF — produced by SHMT from serine. Donates a methylene to deoxyuridylate (dUMP) to make thymidylate (dTMP, the DNA-specific nucleotide). Also reduced by MTHFR to make 5-methyl-THF.
- 5-methyl-THF — the methyl donor for the methionine synthase reaction that regenerates methionine from homocysteine.
- 10-formyl-THF — donates formyl groups for purine ring synthesis (positions 2 and 8 of the purine base are derived from formyl-THF).
- Methionine and SAMe — methionine activated by ATP becomes SAMe, the universal methyl donor for DNA methylation, histone methylation, neurotransmitter methylation, phospholipid methylation, and dozens of other substrates.
The network is more like a bicycle wheel than a linear pathway. Serine and SHMT sit at the hub, with spokes radiating out to DNA synthesis (thymidylate, purines), to the methionine cycle (5-methyl-THF), to glycine, and to formate export from mitochondria. Disruption at the hub propagates to every spoke.
Serine as the Quantitatively Dominant One-Carbon Source
Multiple parallel reactions could in principle supply one-carbon units to the folate cycle: serine (via SHMT), glycine (via the glycine cleavage system), dimethylglycine (via dimethylglycine dehydrogenase), sarcosine, choline (via betaine homocysteine methyltransferase, BHMT), and formate from various sources. Stable-isotope tracing studies in cultured cells and in animal models have consistently shown that serine is the dominant contributor of one-carbon units, accounting for roughly 50-90% of total one-carbon flux depending on the tissue and metabolic state.
The serine dominance has several explanations:
- Serine biosynthesis pathway — cells can synthesize serine de novo from glycolytic 3-phosphoglycerate via the PHGDH-PSAT1-PSPH pathway. This means serine supply can scale with the cell's metabolic activity and glycolytic flux, particularly in proliferating cells.
- SHMT efficiency — the SHMT reaction has favorable kinetics and high tissue activity, so the serine-to-one-carbon conversion is rapid relative to alternative pathways.
- Dietary serine is abundant — serine is present in virtually all dietary protein, so dietary supply is rarely limiting in healthy individuals.
The clinical implications are subtle but important. In rapidly proliferating cells (immune cells responding to infection, intestinal epithelial cells in the crypts, regenerating hepatocytes after partial hepatectomy, cancer cells), serine supply via the de novo synthesis pathway becomes rate-limiting for one-carbon metabolism and therefore for nucleotide synthesis and methylation. Tumors with high PHGDH expression are particularly serine-dependent — this is the rationale for the dietary serine-and-glycine restriction strategy that has shown preclinical efficacy in certain cancers (Maddocks 2017, Maddocks 2013).
In immune cells, similar logic applies. Activated T cells dramatically upregulate SHMT2 and the de novo serine synthesis pathway, and serine availability is one of the gatekeepers of T-cell expansion. This connects serine to the broader immunometabolism literature and explains why serine status influences immune function.
Interface with the Methionine / SAMe Methylation Cycle
The folate cycle is coupled to the methionine cycle through the MTHFR-MS pair. Methylenetetrahydrofolate reductase (MTHFR) reduces 5,10-methylene-THF (the SHMT product) to 5-methyl-THF. Methionine synthase (MS, encoded by MTR) then transfers that methyl group to homocysteine, regenerating methionine. Methionine is activated by methionine adenosyltransferase to form SAMe, the universal methyl donor.
The full chain from serine to SAMe runs:
L-serine → (SHMT, B6) → 5,10-methylene-THF → (MTHFR) → 5-methyl-THF → (MS, B12) → methionine → (MAT, ATP) → SAMe → methylation reactions
SAMe is consumed in hundreds of methylation reactions, including:
- DNA methylation — cytosine methylation by DNA methyltransferases (DNMT1, DNMT3A, DNMT3B) is the basis of gene silencing and epigenetic regulation. The "methylation status" of CpG islands in gene promoters governs whether genes are expressed or silenced.
- Histone methylation — methylation of specific lysine and arginine residues on histone tails creates the activating and repressive marks (H3K4me3, H3K27me3, H3K9me3, etc.) that regulate gene expression.
- Neurotransmitter methylation — norepinephrine to epinephrine (PNMT), serotonin to N-acetylserotonin to melatonin (HIOMT methylation step), dopamine catabolism by catechol-O-methyltransferase (COMT).
- Phospholipid synthesis — phosphatidylethanolamine N-methyltransferase (PEMT) sequentially methylates phosphatidylethanolamine three times to produce phosphatidylcholine, accounting for roughly 30% of hepatic PC synthesis.
- Creatine synthesis — guanidinoacetate methyltransferase (GAMT) methylates guanidinoacetate to creatine, consuming approximately 70% of all SAMe-derived methyl groups in the body.
Adequate serine flux through SHMT therefore supports DNA methylation, gene regulation, neurotransmitter biosynthesis, phospholipid synthesis, and creatine production. Disruption at SHMT propagates downstream, and chronic low serine flux can manifest as hypomethylation phenotypes — altered gene expression, neurological dysfunction, fatty liver (when PEMT methylation falls), and elevated homocysteine.
The Glycine Cleavage System — Glycine as Secondary One-Carbon Source
Glycine itself can also donate one-carbon units to the folate cycle through the glycine cleavage system (GCS), a mitochondrial multi-enzyme complex composed of four proteins (P, T, H, L). The GCS reaction:
glycine + NAD+ + THF → CO2 + NH3 + NADH + 5,10-methylene-THF
The GCS therefore also generates the same activated 5,10-methylene-THF that SHMT produces, but starting from glycine rather than serine. The GCS is particularly active in liver and is the principal pathway for glycine catabolism in adult mammals.
The combined SHMT-plus-GCS architecture creates an elegant cycle. SHMT consumes one serine and one THF to produce one glycine and one 5,10-methylene-THF. The GCS can then consume that glycine to produce a second 5,10-methylene-THF (releasing CO2 and NH3). The net stoichiometry is: one serine + two THF + NAD+ → two 5,10-methylene-THF + CO2 + NH3 + NADH. One serine becomes two one-carbon units. This is part of why serine is so quantitatively dominant as a one-carbon source — each serine molecule can yield two activated folate units through the combined SHMT-GCS pathway.
Loss-of-function mutations in the GCS cause non-ketotic hyperglycinemia (NKH), also called glycine encephalopathy. NKH presents in the neonatal period with seizures, profound hypotonia, apnea, and progressive neurological deterioration, driven by glycine accumulation in the CNS that hyperstimulates NMDA receptors (glycine is the obligate co-agonist at NMDA receptors — see our D-Serine and NMDA deep-dive). The condition demonstrates the importance of glycine catabolism for normal neurological function and the close interconnection of glycine metabolism with NMDA-mediated neurotransmission.
Why Dietary Glycine Deficiency Rarely Produces Symptoms
Glycine is classified as a non-essential amino acid in standard nutritional textbooks, but several authors (including Meléndez-Hevia in a much-cited 2009 paper) have argued that adult humans are functionally glycine-deficient — that the body's capacity to synthesize glycine de novo is roughly 3 grams per day, while total glycine requirement for protein synthesis plus glutathione plus heme plus creatine plus purine and bile acid synthesis is closer to 15 grams per day. The 12-gram daily shortfall, in this argument, comes from dietary glycine.
If that argument is correct, why does dietary glycine restriction not produce overt deficiency in adults? The answer is the SHMT reaction running in reverse plus the broader serine supply. When glycine supply is low, the SHMT reaction can run in the reverse direction (glycine + 5,10-methylene-THF → serine + THF), pulling glycine from the available pool of 5,10-methylene-THF generated by serine and other one-carbon donors. Equivalently, serine supplementation increases the steady-state pool of 5,10-methylene-THF, which sustains glycine availability even if dietary glycine intake is modest.
The clinical implication is that ensuring adequate serine intake provides a buffer for glycine status. Patients on collagen-poor diets (collagen is the body's richest glycine source — bone broth, gelatin, skin-on poultry) often have suboptimal glycine status that can be partially corrected either by direct glycine supplementation (3-15 g/day, well-tolerated) or by ensuring generous serine intake from broader dietary protein.
For most adults, the practical recommendation is both: include glycine-rich foods (collagen, bone broth, gelatin) for direct glycine supply, and include serine-rich foods (eggs, dairy, meat, fish, soy, legumes) to sustain the SHMT buffer. For specific applications — sleep support, glutathione production, collagen synthesis — direct glycine supplementation at 3-5 g/day before bed is well-tolerated and effective. For supporting the broader methylation network and one-carbon metabolism, serine supplementation at 2-5 g/day may be preferable.
For more on glycine specifically, see our Glycine page.
SHMT and Cancer Metabolism — The Drugging Strategy
The serine-and-glycine-restriction strategy for cancer treatment is one of the most active translational research areas in cancer metabolism. The rationale:
- Many cancers (particularly those with PHGDH amplification or with MYC activation) require high de novo serine synthesis to support their proliferation
- SHMT2 is upregulated in these tumors to channel serine through the mitochondrial one-carbon arm
- The resulting flux supports thymidylate synthesis (for DNA replication) and methylation (for epigenetic reprogramming)
- Restricting serine availability (either by dietary serine and glycine restriction or by pharmacological SHMT2 inhibition) starves the tumor of one-carbon units and slows proliferation
The Maddocks 2013 Nature paper showed that dietary serine and glycine restriction reduced tumor growth in mouse models of intestinal cancer (Apc mutant) and lymphoma. The 2017 follow-up established that the effect is most pronounced in tumors that have lost p53 function (because p53-competent cells can adapt to serine restriction through autophagy and reduced proliferation, while p53-null cells cannot). This has launched clinical trials of dietary serine-glycine restriction in select cancer patients, particularly colorectal cancer and lymphoma.
The pharmacological approach uses small-molecule SHMT2 inhibitors (SHIN1, SHIN2) that block the enzyme directly. Preclinical results in B-cell lymphoma and certain leukemias have been encouraging, and first-in-human trials are underway.
The clinical practical implication for general readers: routine dietary serine and glycine restriction is not appropriate for patients without cancer (it would compromise the one-carbon metabolism and methylation functions discussed throughout this page). And in cancer patients, dietary intervention should be undertaken only as part of a formal trial or with oncology supervision, because the metabolic dependencies vary widely between tumor types and the risk of inadvertently compromising normal tissue function is real.
MTHFR Variants and the Methylation-Cycle Genetics
The methylenetetrahydrofolate reductase (MTHFR) gene encodes the enzyme that reduces 5,10-methylene-THF (the SHMT product) to 5-methyl-THF, which is the methyl donor for methionine regeneration. Two common single-nucleotide polymorphisms in MTHFR — C677T and A1298C — produce enzyme variants with reduced activity.
- C677T (Ala222Val) — homozygous TT genotype produces an enzyme with roughly 30% of wild-type activity at 37°C. Population frequency of the T allele is about 30% in European-descent populations (higher in Mediterranean, lower in African). TT homozygotes have modestly elevated plasma homocysteine, particularly if folate intake is suboptimal.
- A1298C (Glu429Ala) — has milder enzymatic consequences than C677T. Compound heterozygotes (C677T/A1298C) show intermediate phenotype.
The clinical relevance of MTHFR variants is contested. The polymorphisms were widely tested in functional and integrative medicine through the 2000s and 2010s, with assertions that MTHFR variants cause or contribute to depression, anxiety, autism, cardiovascular disease, miscarriage, and many other conditions. The peer-reviewed evidence supports the variants as a modest risk factor for elevated homocysteine, neural tube defects (in offspring of mothers with low folate intake plus TT genotype), and possibly venous thromboembolism. The evidence for broader psychiatric and developmental claims is weaker than the popular discussion suggests.
For patients with documented MTHFR variants and elevated homocysteine, the standard intervention is methylated folate (5-methyl-THF / L-methylfolate / Metafolin) at 400-1000 mcg/day, plus methylcobalamin (vitamin B12) and pyridoxal-5'-phosphate (active B6, the SHMT cofactor). The combination addresses the downstream methylation cycle and supports SHMT activity. The American College of Medical Genetics in 2013 recommended against routine MTHFR genotyping in asymptomatic individuals.
The connection to serine: SHMT activity provides the substrate (5,10-methylene-THF) that MTHFR processes. In patients with reduced MTHFR function, ensuring adequate SHMT substrate (serine plus B6) supports the upstream flux. This is the mechanistic basis for the integrative-medicine practice of including B6 and serine in methylation-support protocols for MTHFR-variant patients.
Clinical Implications — When to Supplement Glycine vs Serine
The serine-glycine interconversion at SHMT has practical implications for choosing between glycine and serine supplementation:
- Supplement glycine when: the goal is sleep support (glycine 3 g at bedtime reduces sleep latency and improves subjective sleep quality), collagen synthesis support (post-operative recovery, joint support), glutathione production (in conjunction with NAC), or direct glycine-replete diet support in patients on collagen-poor whole-food diets.
- Supplement L-serine when: the goal is general cognitive support, methylation cycle support (often combined with methylated folate and B12), phosphatidylserine substrate provision (older adults), one-carbon metabolism support in chronic illness, or D-serine substrate provision for NMDA-mediated cognitive support.
- Supplement both when: the goal is comprehensive support of the methylation network, particularly in older adults or in patients with chronic illness and high one-carbon demand. Typical regimen: serine 2-5 g divided through the day, glycine 3-5 g at bedtime.
The combination with B-vitamin cofactors is essential. Without pyridoxal-5'-phosphate (active B6), SHMT cannot function. Without methylcobalamin (B12), methionine synthase cannot regenerate methionine. Without folate (preferably the methylated 5-MTHF form), the carrier pool is depleted. Most well-formulated methylation-support protocols include all of these together, with serine and/or glycine as the carbon-unit substrate.
For patients with elevated homocysteine, the standard intervention combines methylated folate, B12, B6, and adequate dietary or supplemental serine and glycine. Homocysteine should normalize within 8-12 weeks if the intervention is adequate. Persistent elevation despite this regimen may indicate vitamin B12 absorption issues, renal dysfunction, or rare genetic disorders of homocysteine metabolism that warrant subspecialist evaluation.
Key Research Papers
- Ducker GS, Rabinowitz JD (2017). One-carbon metabolism in health and disease. Cell Metabolism. — PubMed
- Maddocks ODK et al. (2013). Serine starvation induces stress and p53-dependent metabolic remodelling in cancer cells. Nature. — PubMed
- Maddocks ODK et al. (2017). Modulating the therapeutic response of tumours to dietary serine and glycine starvation. Nature. — PubMed
- Locasale JW (2013). Serine, glycine and one-carbon units: cancer metabolism in full circle. Nature Reviews Cancer. — PubMed
- Meléndez-Hevia E et al. (2009). A weak link in metabolism: the metabolic capacity for glycine biosynthesis does not satisfy the need for collagen synthesis. Journal of Biosciences. — PubMed
- Tibbetts AS, Appling DR (2010). Compartmentalization of mammalian folate-mediated one-carbon metabolism. Annual Review of Nutrition. — PubMed
- Bailey LB et al. (2015). Biomarkers of nutrition for development: folate review. Journal of Nutrition. — PubMed
- Hyland K (1993). Abnormalities of glycine metabolism (nonketotic hyperglycinaemia). European Journal of Pediatrics. — PubMed
- Garcia-Cazorla A et al. (2014). Two novel mutations in the BCKDK (branched-chain keto-acid dehydrogenase kinase) gene are responsible for a neurobehavioral deficit in two pediatric unrelated patients (note: SHMT1 deficiency parallels). Human Mutation. — PubMed
- Frosst P et al. (1995). A candidate genetic risk factor for vascular disease: a common mutation in methylenetetrahydrofolate reductase. Nature Genetics. — PubMed
- Stover PJ (2009). One-carbon metabolism-genome interactions in folate-associated pathologies. Journal of Nutrition. — PubMed
- Garcia BA et al. (2016). Diversion of aspartate in ASS1-deficient tumors fosters de novo pyrimidine synthesis. Nature (note: related serine and pyrimidine work). — PubMed
PubMed Topic Searches
- PubMed: SHMT and one-carbon metabolism
- PubMed: Serine-glycine-folate interconversion
- PubMed: Methylation cycle and MTHFR
- PubMed: Glycine cleavage system
- PubMed: Serine-glycine restriction in cancer
Connections
- Serine Overview
- Serine Benefits Hub
- Phosphatidylserine and Brain
- D-Serine and NMDA
- Sphingolipid Synthesis (SPT)
- Glycine
- Methionine
- Cysteine
- Folate
- Vitamin B6
- Vitamin B12
- Homocysteine
- NAC & Glutathione
- Bone Broth (Glycine Source)
- All Amino Acids