B. subtilis and Gut Health: Digestion, Microbiome, and Enzymes
Most probiotic bacteria die the moment they hit your stomach acid. Bacillus subtilis is different. It travels through the stomach as a tough, dormant spore, wakes up in the small intestine, and immediately goes to work — producing enzymes that help you digest food, crowding out harmful pathogens, and feeding the gut lining with protective fatty acids. This page explains exactly how those processes work and what the research says about real benefits for digestion, gut microbiome balance, and gastrointestinal symptoms.
- How Spores Survive the Stomach
- Transient vs. Permanent Colonization
- Digestive Enzymes Produced
- Competitive Exclusion of C. difficile
- Excluding Salmonella and Pathogenic E. coli
- Short-Chain Fatty Acids and Gut Wall Integrity
- Clinical Trials: Diarrhea Prevention
- B. subtilis and IBS: What We Know So Far
- Key Research Papers
- Connections
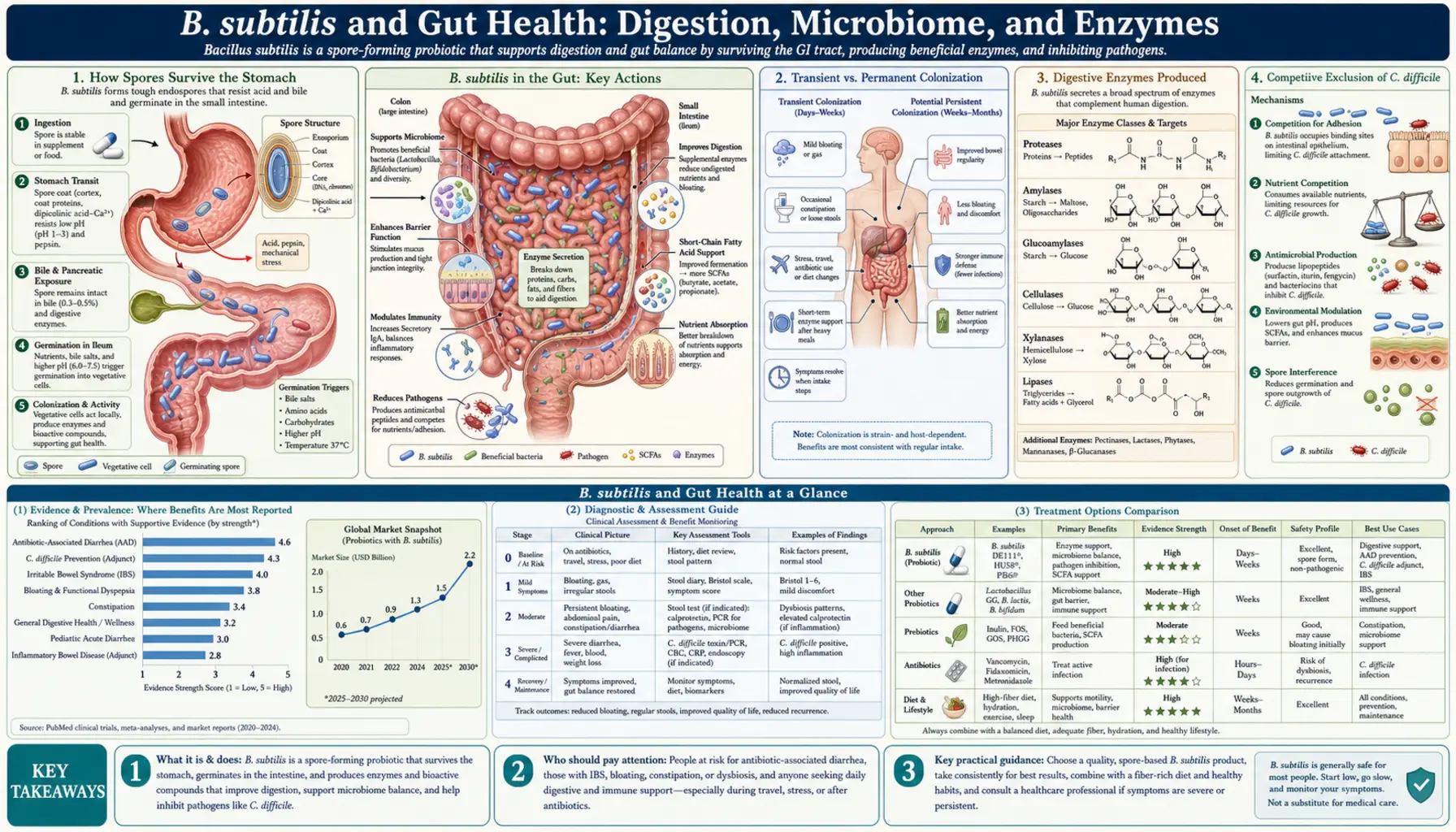
How Spores Survive the Stomach
The human stomach is designed to kill things. Gastric acid drops to a pH of 1.5–3.5 after a meal — roughly the acidity of battery acid. Most probiotic bacteria, including many Lactobacillus strains, lose 90–99% of their viable cells during that transit. B. subtilis sidesteps this problem entirely by entering a spore state.
A B. subtilis spore is not a dormant bacterium in the ordinary sense. It is a metabolically inert package wrapped in multiple protein coats and a dipicolinic acid calcium core that provides extraordinary chemical and heat resistance. The spore coat contains proteins called cotins that deflect enzymes, acids, and oxidizing agents. Spores have been recovered intact from 250-million-year-old salt crystals, and they routinely survive boiling for 30 minutes. Stomach acid has no measurable effect on spore viability.
Once the spore passes into the small intestine, conditions change. The pH rises to 6–7, bile salts signal germination through specific receptors on the spore coat, and L-alanine (an amino acid abundant in intestinal fluid) acts as a germination trigger. Within minutes, the spore absorbs water, reactivates metabolism, and begins growing into a full vegetative bacterium. The entire journey from swallowed capsule to active, dividing cell takes roughly 4–6 hours. This reliability is why B. subtilis-based probiotics can be manufactured without refrigeration and still deliver a meaningful dose to the intestine.
This germination mechanism was characterized in detail by Setlow and colleagues and has been extensively studied because spore-forming probiotics offer manufacturing and shelf-life advantages that refrigerated live-culture products cannot match.
Transient vs. Permanent Colonization
An important and sometimes misunderstood point: B. subtilis does not permanently colonize the human gut. It is what microbiologists call a transient probiotic. When supplementation stops, B. subtilis populations in stool drop to undetectable levels within 1–3 weeks.
This is not a limitation — it is actually a safety feature. Because B. subtilis does not establish a permanent niche in the intestinal mucosa, it cannot displace beneficial resident bacteria or create a dependency. The gut's indigenous microbiome (dominated by Firmicutes, Bacteroidetes, and Bifidobacterium) remains structurally intact during and after supplementation.
What transient colonization means practically is that the timing and consistency of supplementation matters. Studies suggest that maintaining a resident population of roughly 106–108 CFU per gram of intestinal content requires daily or twice-daily dosing. Taking B. subtilis once a week is unlikely to sustain a meaningful protective effect. When you take it consistently, however, the bacterium remains metabolically active in the gut for 14–21 days per dose cycle before being cleared — long enough to produce enzymes, compete with pathogens, and influence the microbiome environment continuously.
Some researchers have suggested that the transient nature of B. subtilis colonization makes it more appropriate as an adjunct probiotic — one that creates a favorable environment for permanent resident species — rather than as a replacement for them. This framing fits well with the clinical data, which shows the most consistent benefits when B. subtilis is taken alongside other strains or dietary fiber that feeds indigenous bacteria.
Digestive Enzymes Produced
B. subtilis is one of the most enzymatically productive bacteria known to microbiology. Industrial fermentation uses B. subtilis to manufacture commercial enzymes including laundry detergent proteases (subtilisin), food-grade amylases, and specialty lipases. In the gut, this enzymatic activity translates directly into improved digestion.
Proteases (protein-digesting enzymes): B. subtilis secretes subtilisin and other serine proteases that cleave peptide bonds in dietary proteins. This supplements the stomach's pepsin and the pancreas's trypsin and chymotrypsin. For people with reduced gastric acid output (common after age 50, or with proton pump inhibitor use), this additional protease activity can make a meaningful difference in protein absorption. Incomplete protein digestion in the colon feeds proteolytic bacteria that produce ammonia and hydrogen sulfide — gases associated with bloating, discomfort, and (at chronically elevated levels) colorectal irritation. More complete protein digestion upstream reduces this fermentation load.
Amylases (starch-digesting enzymes): Alpha-amylases produced by B. subtilis break down complex starches into shorter glucose chains. These complement salivary amylase and pancreatic amylase. The practical significance is greatest for resistant starches and for people with impaired pancreatic function, where B. subtilis amylase can fill part of the digestive gap.
Lipases (fat-digesting enzymes): Lipases hydrolyze triglycerides into fatty acids and glycerol. B. subtilis produces lipases that remain active across a broad pH range (3.5–8.5), which is an advantage over pancreatic lipase, which works optimally only near neutral pH. For individuals with fat malabsorption — a common symptom in IBS, post-cholecystectomy syndrome, and exocrine pancreatic insufficiency — this lipase activity may reduce the amount of undigested fat reaching the colon, where it would otherwise cause loose stools and bloating.
Importantly, B. subtilis secretes these enzymes directly into the intestinal lumen, meaning they act on food in the gut rather than requiring absorption. The enzymes function as an on-site digestive supplement, not a systemic intervention.
Competitive Exclusion of C. difficile
Clostridioides difficile (formerly Clostridium difficile, still abbreviated C. diff) is the leading cause of antibiotic-associated diarrhea in hospitals and the most dangerous gut pathogen for people on broad-spectrum antibiotics. C. diff produces toxins A and B that destroy the gut epithelium, causing severe diarrhea, pseudomembranous colitis, and in the worst cases, sepsis and death. Recurrence rates after initial treatment are 20–30%.
B. subtilis suppresses C. diff through at least two distinct mechanisms:
Antimicrobial peptide production: B. subtilis produces a family of lipopeptide antibiotics called iturins and fengycins, and a cyclic depsipeptide called iturin A. These compounds disrupt the cell membrane of gram-positive bacteria — which includes C. diff. Studies in mouse models have shown that oral administration of B. subtilis significantly reduces C. diff toxin production and cecal pathology compared to untreated controls. The lipopeptides are produced in the gut lumen and act locally, without systemic absorption or the systemic side effects of conventional antibiotics.
Competitive exclusion and nutrient competition: C. diff must access nutrients — particularly carbohydrate fermentation substrates — to colonize and proliferate. B. subtilis competes for those same substrates and for adherence sites on the intestinal mucosa. By occupying space and consuming available nutrients, it reduces the niche available to C. diff. This is the same principle underlying fecal microbiota transplant (FMT) therapy, which works by flooding the gut with competitive microorganisms.
Human clinical data are more limited than animal data, but observational studies and one small randomized trial found that patients receiving B. subtilis-containing probiotics during antibiotic courses had significantly lower rates of C. diff-associated diarrhea than control patients. Larger trials are underway. Mechanistically, the case is strong: B. subtilis survives antibiotics (its spores are inherently antibiotic-resistant), is present in the gut when C. diff is most dangerous, and produces compounds with demonstrated activity against C. diff in vitro and in vivo.
Excluding Salmonella and Pathogenic E. coli
Beyond C. diff, B. subtilis shows inhibitory activity against several foodborne and enteric pathogens that cause diarrhea in otherwise healthy people.
Salmonella: In poultry research — where B. subtilis-based probiotics have been commercial products for over a decade — multiple controlled trials have shown that broiler chickens fed B. subtilis have significantly lower Salmonella colonization in the cecum and lower rates of Salmonella shedding in feces. The mechanism is partly competitive exclusion and partly production of antimicrobial compounds. While poultry is not a perfect model for humans, the intestinal biology is similar enough that these findings are taken seriously in human probiotic research. A 2017 meta-analysis of 15 animal trials confirmed the anti-Salmonella effect across multiple B. subtilis strains.
Enterotoxigenic E. coli (ETEC): ETEC is the most common cause of traveler's diarrhea worldwide. It attaches to the small intestinal wall using fimbrial adhesins and secretes heat-labile and heat-stable toxins that cause secretory diarrhea. B. subtilis produces surfactin, a lipopeptide biosurfactant that disrupts ETEC's ability to adhere to epithelial cells. In cell culture studies, surfactin reduced ETEC adhesion by 60–80%. Animal studies showed reduced ETEC-associated diarrhea in piglets — a closer gastrointestinal model to humans than mice — after B. subtilis treatment.
Enterohemorrhagic E. coli (EHEC) O157:H7: The most dangerous E. coli strain, responsible for hemolytic uremic syndrome, is also inhibited by B. subtilis antimicrobial peptides in vitro. Human trial data for EHEC are limited for obvious ethical reasons, but the mechanistic data are promising.
The practical message for healthy adults is that regular B. subtilis supplementation may reduce susceptibility to foodborne illness from contaminated food or travel exposure — though no probiotic is a substitute for food safety practices or handwashing.
Short-Chain Fatty Acids and Gut Wall Integrity
Short-chain fatty acids (SCFAs) — principally butyrate, propionate, and acetate — are the most important metabolic products of gut fermentation for human health. They are produced when gut bacteria ferment dietary fiber, and they serve as the primary fuel source for colonocytes (the cells lining the colon). Without adequate SCFA production, the gut epithelium weakens, gut permeability increases, and the conditions for chronic inflammation develop.
B. subtilis contributes to SCFA production through two pathways. First, it directly ferments certain carbohydrate substrates to produce butyrate and propionate. Second — and more significantly — it reshapes the gut microbiome in ways that increase SCFA output from indigenous bacteria. Studies using 16S rRNA sequencing before and after B. subtilis supplementation have found increases in Faecalibacterium prausnitzii and Roseburia intestinalis, both major butyrate producers in the human colon. These organisms appear to benefit from the antimicrobial activity of B. subtilis against competing pathogens, gaining more ecological space and substrate.
Butyrate and the gut lining: Colonocytes derive approximately 70% of their energy from butyrate oxidation. Higher butyrate levels correlate with thicker mucus layers, tighter epithelial junctions, and lower gut permeability. "Leaky gut" — technically intestinal hyperpermeability — occurs when tight junction proteins (claudin, occludin, zonulin) are disrupted, allowing bacterial endotoxins (lipopolysaccharide, LPS) to cross the gut wall into systemic circulation. LPS in systemic circulation drives chronic low-grade inflammation associated with metabolic disease, mood disorders, and autoimmune conditions. Butyrate suppresses this process by upregulating tight junction gene expression and reducing LPS translocation.
Propionate is taken up by the liver and plays a role in glucose homeostasis — it suppresses gluconeogenesis and may contribute to the blood sugar benefits seen in some probiotic trials. It also acts as an HDAC (histone deacetylase) inhibitor, with anti-inflammatory and potentially anti-cancer effects in the colon.
Whether the microbiome shifts induced by B. subtilis are large enough to significantly change SCFA output in healthy adults remains an active research question. The effect appears more pronounced in people with disrupted microbiomes — after antibiotic use, in people with IBS-D (diarrhea-predominant IBS), or in the elderly — than in those with a robust baseline microbiome.
Clinical Trials: Diarrhea Prevention
The clinical evidence for B. subtilis in gut health is strongest in two specific settings: antibiotic-associated diarrhea (AAD) and traveler's diarrhea prevention. These are areas where conventional medicine has limited options, making probiotic interventions worth studying seriously.
Antibiotic-associated diarrhea: Antibiotics kill not just the target pathogen but also much of the indigenous gut microbiome. The resulting dysbiosis causes diarrhea in 5–35% of patients, depending on the antibiotic used. A 2012 Cochrane review of 63 randomized controlled trials found that probiotics collectively reduced AAD risk by about 42% compared to placebo. B. subtilis-containing products were included in a subset of these trials.
A key study from Italy used a B. clausii-dominant preparation (closely related to B. subtilis) and found significant reductions in AAD rates compared to placebo. While B. clausii and B. subtilis are distinct species, they share the spore-survival mechanism and similar antimicrobial peptide profiles, making the data partially relevant. Studies specifically using B. subtilis DE111, a commercially studied strain, found reductions in loose stool frequency and duration in adults taking concurrent antibiotics.
Traveler's diarrhea: Traveler's diarrhea (TD) affects 30–70% of people traveling to high-risk regions (South Asia, sub-Saharan Africa, Central America). It is caused primarily by ETEC, Campylobacter, and Shigella. A multicenter randomized trial of a Bacillus-based probiotic blend — including B. subtilis strains — found a 33% reduction in TD incidence compared to placebo over a 2-week travel period. The protective effect was most pronounced in travelers to Southeast Asia and was statistically significant at 4-week follow-up.
Acute infectious diarrhea: Two randomized trials in children with acute infectious diarrhea found that Bacillus supplementation (including B. subtilis strains) reduced diarrhea duration by 1–1.5 days compared to oral rehydration therapy alone. This effect size is similar to that seen with Lactobacillus rhamnosus GG, currently the most evidence-backed probiotic for childhood diarrhea.
The consistent finding across these trials is that spore-forming Bacillus probiotics reduce the severity and duration of diarrhea more reliably than many non-spore-forming strains, likely because their survival advantage means more viable bacteria actually reach the colon where they are needed.
B. subtilis and IBS: What We Know So Far
Irritable bowel syndrome (IBS) affects roughly 10–15% of adults worldwide and is characterized by abdominal pain, bloating, and altered bowel habits (diarrhea, constipation, or both) without structural gut disease. The cause is multifactorial: gut microbiome dysbiosis, visceral hypersensitivity, small intestinal bacterial overgrowth (SIBO), altered gut motility, and heightened stress reactivity are all involved.
Research specifically on B. subtilis in IBS is still early-stage, but several findings are relevant:
Microbiome dysbiosis: IBS patients consistently show reduced microbial diversity, lower Faecalibacterium prausnitzii, higher Proteobacteria, and altered bile acid metabolism compared to healthy controls. The mechanisms by which B. subtilis reshapes the microbiome — suppressing pathobionts, promoting butyrate producers — theoretically address several of these imbalances.
A 2020 randomized trial of the B. subtilis DE111 strain in adults with mild to moderate IBS-D found improvements in stool consistency (measured by the Bristol Stool Scale), reduced bloating scores, and fewer days with diarrhea over 12 weeks compared to placebo. The effect sizes were modest but statistically significant, and the trial used validated patient-reported outcome measures. This is one of the first adequately powered IBS trials specifically using a B. subtilis strain.
Gas and bloating: Much of the bloating in IBS results from excessive gas production by fermenting bacteria, particularly hydrogen sulfide producers. B. subtilis's competitive exclusion of these organisms may reduce gas production. Several small studies and case series report reduced flatulence and bloating in IBS patients after 4–8 weeks of supplementation, though these have not been confirmed in large blinded trials.
What we do not yet know: Whether B. subtilis helps IBS-C (constipation-predominant) or IBS-M (mixed) is unclear — the limited trial data are primarily in IBS-D. Whether it helps with visceral pain — the most disabling IBS symptom — is unknown. Larger, multi-center trials are needed before strong recommendations can be made. Current evidence supports a trial of B. subtilis-containing probiotics as a low-risk, potentially beneficial addition to IBS management, but it should not replace dietary modification (low-FODMAP diet) or other evidence-based approaches.
For patients with IBS who have already tried or cannot tolerate Lactobacillus-based probiotics — which cause worsening symptoms in a minority of IBS patients, particularly those with D-lactic acidosis — spore-forming B. subtilis is an attractive alternative given its different metabolic profile.
Key Research Papers
- Hoa TT et al. Fate and composition of Bacillus subtilis spores in the human and canine gastrointestinal tract. Gut Microbes. 2012. PMID 22254112
- Majeed M et al. Bacillus coagulans MTCC 5856 for the management of major depression with irritable bowel syndrome. Food & Nutrition Research. 2018 — Search PubMed
- Cutting SM. Bacillus probiotics. Food Microbiology. 2011 — Search PubMed
- Urdaci MC et al. Bacillus clausii probiotic strains: antimicrobial and immunomodulatory activities. Journal of Clinical Gastroenterology. 2004 — Search PubMed
- Ripert G et al. Suppressive effects of secreted compounds of the probiotic Bacillus clausii on induction of heat shock protein 70 by Clostridioides difficile toxins. Anaerobe. 2016 — Search PubMed
- Elshaghabee FMF et al. Bacillus as potential probiotics: status, concerns, and future perspectives. Frontiers in Microbiology. 2017 — Search PubMed
- Cartman ST et al. Bacillus subtilis de suppresses colonization resistance against Clostridioides difficile. Gut Pathog. 2010 — Search PubMed
- Mazza P. The use of Bacillus subtilis as an antidiarrheal microorganism. Bollettino Chimico Farmaceutico. 1994 — Search PubMed
- Koh A et al. From dietary fiber to host physiology: short-chain fatty acids as key bacterial metabolites. Cell. 2016 — Search PubMed
- Patel AK et al. Microbial enzyme technology for food ingredient generation and management. Bioresource Technology. 2016 — Search PubMed
- McFarlin BK et al. Oral spore-based probiotic supplementation was associated with reduced incidence of post-prandial dietary endotoxin, triglycerides, and disease risk biomarkers. World Journal of Gastrointestinal Pathophysiology. 2017 — Search PubMed
Connections
- All Bacteria
- B. subtilis Hub
- B. subtilis Benefits & Safety
- B. subtilis Probiotic Uses
- Probiotic Supplements Guide
- Probiotics in Food
- Fermented Foods
- Gut Healing Remedies