Hepatitis: History and Discovery
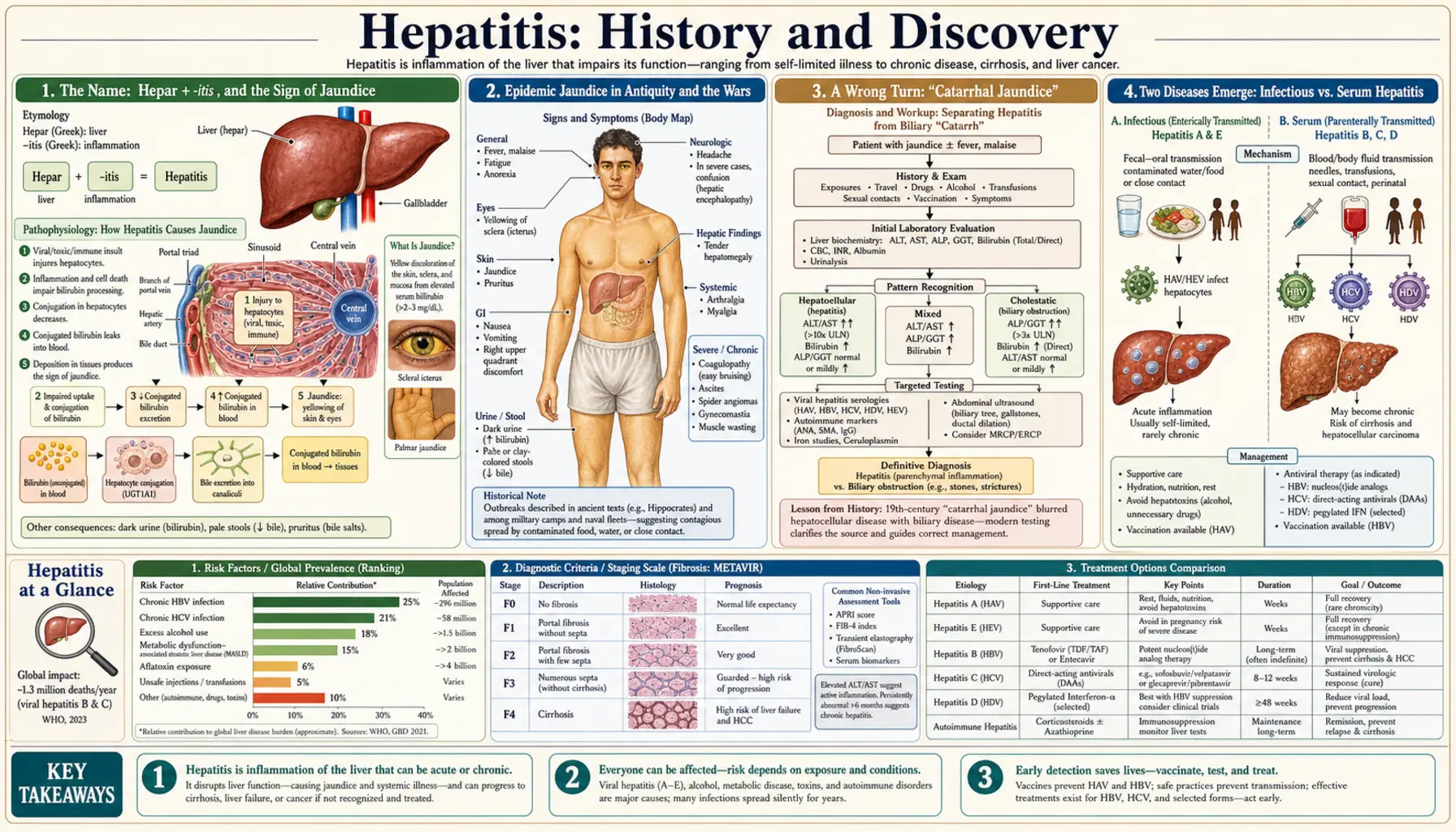
The word hepatitis simply means “inflammation of the liver,” from the Greek hepar (liver) and the suffix -itis (inflammation). The visible sign of liver trouble — the yellow skin and eyes of jaundice — has been recognized since antiquity: Hippocrates described an “epidemic jaundice” around 400 BCE, and outbreaks of “campaign jaundice” shadowed armies through almost every major war into the twentieth century. But the causes — a family of unrelated viruses that all happen to attack the same organ — remained hidden until modern virology. The breakthroughs came in a rush: Baruch Blumberg found the “Australia antigen” (hepatitis B) in the 1960s and won a Nobel Prize in 1976; hepatitis A was caught under the electron microscope in 1973; and the elusive virus behind transfusion hepatitis was finally cloned in 1989 and named hepatitis C — work honored with a second Nobel Prize in 2020. This page traces that long road, carefully separating the ancient clinical syndrome of jaundice from the modern discovery of the distinct viruses, and flags older ideas that turned out to be wrong.
Table of Contents
- The Name: Hepar + -itis, and the Sign of Jaundice
- Epidemic Jaundice in Antiquity and the Wars
- A Wrong Turn: “Catarrhal Jaundice”
- Two Diseases Emerge: Infectious vs. Serum Hepatitis
- Blumberg and the Australia Antigen (Hepatitis B)
- Catching Hepatitis A, 1973
- “Non-A, Non-B” and the Discovery of Hepatitis C
- Hepatitis D and Hepatitis E
- Vaccines, Cures, and the Modern Legacy
- Research Papers and References
- Connections
- Featured Videos
The Name: Hepar + -itis, and the Sign of Jaundice
The term hepatitis is built from two ancient Greek elements: hēpar (genitive hēpatos), meaning “liver,” and the medical suffix -itis, meaning “inflammation of.” The same root gives us hepatic (relating to the liver), hepatology (its study), and hepatocyte (a liver cell). Literally, then, “hepatitis” describes a state — an inflamed liver — and says nothing at all about the cause. That neutrality is important to the history that follows: for most of recorded time, people could see the effects of liver inflammation long before anyone could name what produced it.
The most dramatic visible effect is jaundice (also called icterus), the yellowing of the skin and the whites of the eyes that appears when a diseased liver can no longer clear the yellow-brown pigment bilirubin from the blood. Because jaundice is so striking and so easy to recognize, the early history of liver disease is really a history of jaundice — a symptom shared by many different conditions. Some of those yellow patients had what we would now call viral hepatitis; others had gallstones, malaria, toxins, or other liver injuries entirely. Untangling that single visible sign into its many hidden causes took more than two thousand years and the invention of virology, immunology, and molecular biology.
So when older texts speak of “epidemic jaundice” or “catarrhal jaundice,” we should read those as descriptions of a clinical syndrome — people turning yellow, often in clusters — rather than as a diagnosis of any specific virus. The modern, virus-by-virus picture (hepatitis A through E) is a twentieth-century achievement. The rest of this article keeps that distinction in view, because conflating the ancient symptom with the modern microbe is one of the commonest mistakes in popular medical history.
Epidemic Jaundice in Antiquity and the Wars
Outbreaks of jaundice were noticed in the ancient world. The Greek physician Hippocrates, writing around 400 BCE, is widely credited with describing an “epidemic jaundice” — a contagious-seeming yellowing that spread through a population — among the many conditions catalogued in the Hippocratic writings. Historians of medicine caution that we cannot say which modern disease this was: an epidemic of jaundice in antiquity could plausibly have been hepatitis A or hepatitis E (both spread by contaminated food and water) or yet another cause. The honest summary is that the ancients recognized a transmissible jaundice syndrome, not that they discovered any particular hepatitis virus. These classical observations are named here as historical sources, not as modern citations.
From antiquity onward, the most reliable trigger for large jaundice epidemics was war. Crowded, unsanitary camps with fouled water are an ideal setting for the fecal-oral spread of what we now call hepatitis A and E, and chroniclers recorded waves of jaundice in military campaigns for centuries — so regularly that the condition earned the names “campaign jaundice” and “epidemic jaundice.” Major outbreaks were reported in conflicts ranging from the American Civil War to the Gallipoli campaign of 1915, and hepatitis heavily afflicted the armies of the First World War and again the German forces on the Eastern Front in the Second World War. For armies, jaundice was a serious cause of lost manpower long before its viral nature was understood.
This military pattern is more than a curiosity: it was the great wartime epidemics of the twentieth century, and especially the jaundice that followed certain vaccinations given to soldiers, that finally pushed researchers to realize that “jaundice” was not one disease but at least two, transmitted in two very different ways. The clinical observations of antiquity and the battlefield, in other words, set the stage; the laboratory revolution that decoded them did not begin in earnest until the 1940s.
A Wrong Turn: “Catarrhal Jaundice”
Before the viral story could be written, nineteenth-century medicine took a long detour down a mistaken explanation, and it is worth telling because it shows how a plausible-sounding hypothesis can dominate a field for decades. In 1865 the great German pathologist Rudolf Virchow examined a patient who had died of epidemic jaundice and found the lower end of the common bile duct plugged with a mass of mucus. Reasoning from that single finding, he proposed that the disease was caused by catarrh — an inflammatory build-up of mucus — physically blocking the bile duct so that bile backed up and stained the body yellow. The condition was accordingly christened “catarrhal jaundice.”
It is important to label this clearly as a hypothesis that turned out to be wrong. Virchow’s mucous-plug theory treated hepatitis as a mechanical plumbing problem in the bile duct rather than as an infection of the liver cells themselves. Yet because Virchow was the most authoritative pathologist of his age, the catarrhal-jaundice idea was widely accepted and taught well into the twentieth century — many clinicians clung to it right up to the Second World War. It delayed recognition that the real damage was happening inside the liver, caused by an infectious agent, not by a blob of mucus at the duct’s opening.
The catarrhal-jaundice era is a useful reminder of how scientific understanding actually advances: not in a straight line, but through hypotheses that are proposed, accepted, and eventually overturned by better evidence. Only when the wartime studies of the 1930s and 1940s showed that jaundice could be transmitted from person to person — and in two distinct patterns — did the mucous-plug explanation finally collapse, clearing the way for the modern viral framework.
Two Diseases Emerge: Infectious vs. Serum Hepatitis
The first hard clue that more than one disease hid behind “jaundice” came from medicine’s own needles. As early as 1885, the German physician Lurman investigated an outbreak in Bremen in which shipyard and dock workers, vaccinated against smallpox using human-derived lymph, developed jaundice weeks to months later — while workers given a different lymph batch did not. This is generally regarded as the first clearly documented epidemic of what would later be called serum hepatitis: a jaundice spread not by food and water but by blood and blood products. Similar “post-inoculation” and “post-transfusion” jaundice was noted repeatedly over the following decades.
The decisive evidence came during and after the Second World War. Enormous outbreaks of jaundice followed certain batches of yellow-fever vaccine that had been stabilized with human serum, sickening tens of thousands of soldiers, while other jaundice continued to spread the old fecal-oral way through contaminated food and water. Careful human-volunteer transmission studies in the 1930s and 1940s established that there were two hepatitis diseases with different incubation periods and different routes: a short-incubation illness spread by the fecal-oral route, and a longer-incubation illness spread by blood. Crucially, recovering from one did not protect against the other, proving they were caused by different agents.
In 1947 the British virologist F. O. MacCallum proposed the clean terminology that organized this insight and is still used today: hepatitis A for the contagious, fecal-orally spread “infectious hepatitis,” and hepatitis B for the blood-borne “serum hepatitis.” (MacCallum’s own role traced back to noticing that British soldiers developed hepatitis some months after receiving a yellow-fever vaccine.) A landmark American effort of the same era was Saul Krugman’s work at the Willowbrook State School, where studies of two transmissible agents he labelled MS-1 and MS-2 helped confirm that hepatitis A and hepatitis B were genuinely distinct — though those institutional studies, conducted on intellectually disabled children, are now also remembered as a serious cautionary tale in research ethics. By the late 1940s, then, medicine had two named diseases but still had not seen either virus. That final step belonged to the next two decades.
Blumberg and the Australia Antigen (Hepatitis B)
The modern era of hepatitis virology opened almost by accident, and from an unexpected direction. The American physician and geneticist Baruch S. Blumberg was not hunting for a hepatitis virus at all; he was studying inherited variation in human blood proteins across populations. While testing the sera of patients who had received many transfusions against blood samples from people around the world, he and his colleagues found, in 1963, an antibody in a New York hemophilia patient that reacted with an antigen in the blood of an Australian Aboriginal man. They named the mystery protein the “Australia antigen.” The first report appeared in 1965 (Blumberg, Alter, and Visnich, in JAMA), where the antigen was initially — and wrongly — suspected of being linked to leukemia.
Over the next few years the true meaning of the Australia antigen came into focus. By 1967, accumulating evidence showed that the antigen was strongly associated with viral hepatitis — it was, in fact, a piece of the outer coat of the hepatitis B virus, the protein now known as the hepatitis B surface antigen (HBsAg). In 1970, working in London, David Dane used electron microscopy to visualize the complete hepatitis B virus particle, still called the Dane particle in his honor. For the first time, one of the hepatitis agents had a face, a marker, and a name.
The practical consequences were immense. Because HBsAg could be detected in blood, donated blood could now be screened to keep hepatitis B out of the transfusion supply — a change that prevented countless infections. The same surface antigen later became the basis of a vaccine. For these discoveries “concerning new mechanisms for the origin and dissemination of infectious diseases,” Blumberg shared the 1976 Nobel Prize in Physiology or Medicine (with D. Carleton Gajdusek, recognized for separate work on slow viral infections). Blumberg’s serendipitous discovery is the hinge on which the whole modern history of hepatitis turns: it converted a vague, ancient symptom into a specific, testable, ultimately preventable viral infection.
Catching Hepatitis A, 1973
With hepatitis B’s marker in hand, attention turned to the other classic agent — the fecal-orally transmitted “infectious hepatitis,” hepatitis A — which had been named by MacCallum in 1947 but never actually seen. The breakthrough came in 1973, when Stephen M. Feinstone, working with Albert Z. Kapikian and Robert H. Purcell at the U.S. National Institutes of Health, applied a clever technique called immune electron microscopy. By mixing stool from patients in the acute phase of hepatitis A with antibody-rich serum from recovered patients, they clumped together and made visible tiny 27-nanometer virus particles — the hepatitis A virus (HAV) itself.
Their landmark report, “Hepatitis A: Detection by Immune Electron Microscopy of a Viruslike Antigen Associated with Acute Illness,” was published in Science in 1973 (volume 182, pages 1026–1028). The evidence was compelling because every hepatitis A patient tested mounted an antibody response to the particle, exactly as expected for the true cause of the disease. This is a clean example of how, once one virus (hepatitis B) had been pinned down, the tools and confidence existed to capture the next.
Identifying HAV opened the door to its own diagnostic blood tests and, eventually, to an effective inactivated hepatitis A vaccine, licensed in the 1990s. Just as importantly, isolating both hepatitis A and hepatitis B sharpened a frustrating new problem: a large share of hepatitis cases — especially those following blood transfusion — tested negative for both viruses. Something else was clearly out there. Physicians gave that residual category a deliberately humble, placeholder name: “non-A, non-B hepatitis.”
“Non-A, Non-B” and the Discovery of Hepatitis C
The hunt for the cause of non-A, non-B hepatitis became one of the great detective stories of late-twentieth-century medicine. In the 1970s, working at the NIH, Harvey J. Alter and colleagues showed that much of the hepatitis still occurring after blood transfusions was caused by neither hepatitis A nor hepatitis B, and that this mysterious agent could be transmitted to chimpanzees — proving a specific infectious cause existed even though no one could see or name it. For more than a decade the agent resisted every classical method of virology: it could not be grown in culture or visualized, and it stayed stubbornly anonymous.
The decisive blow was struck with the new tools of molecular biology. At the biotechnology company Chiron, Michael Houghton together with Qui-Lim Choo and George Kuo — using infected chimpanzee material supplied by Daniel W. Bradley of the CDC — spent years building and screening DNA libraries until, in 1989, they fished out a single viral clone from the blood of an infected animal. They had cloned the genome of a previously unknown virus directly, without ever seeing it under a microscope, and named it the hepatitis C virus (HCV). Two landmark papers in Science in 1989 announced both the cloned genome and a blood test for antibodies to it. That antibody test, deployed for blood-bank screening, swiftly and dramatically reduced transfusion-transmitted hepatitis worldwide.
One piece of proof remained: showing that this cloned virus, on its own, actually caused disease. That came from Charles M. Rice, who engineered a complete, functional copy of the HCV genome and demonstrated that it could replicate and produce hepatitis in chimpanzees — closing the logical loop and confirming HCV as the true agent. For this collective achievement, the 2020 Nobel Prize in Physiology or Medicine was awarded to Harvey J. Alter, Michael Houghton, and Charles M. Rice “for the discovery of the Hepatitis C virus.” (The Nobel committee’s choice of three names from a larger team that included Choo, Kuo, and Bradley was widely noted and debated, a reminder that landmark discoveries are usually the work of many hands.) From a placeholder called “non-A, non-B” to a Nobel Prize took just over thirty years.
Hepatitis D and Hepatitis E
The familiar alphabet of hepatitis does not stop at C. In 1977, in Turin, the Italian physician Mario Rizzetto noticed an unusual new antigen — which he called the delta antigen — in the liver-cell nuclei of some patients who were already infected with hepatitis B and had unusually severe disease. This turned out to be a remarkable and unique agent: the hepatitis D virus (HDV), sometimes called the delta agent, a small, defective virus that cannot replicate on its own. It is a true biological parasite of hepatitis B, borrowing the hepatitis B surface antigen as its own envelope, so that only people infected with hepatitis B can be infected with hepatitis D. Co-infection tends to make liver disease markedly worse, which is one more reason the hepatitis B vaccine is so valuable — preventing B also prevents D.
The last of the classic five, hepatitis E (HEV), accounts for much of the old “enterically transmitted non-A, non-B hepatitis” — epidemic, waterborne jaundice resembling hepatitis A and historically common where sanitation is poor. Its existence was confirmed in a now-famous act of scientific self-experimentation: in 1983 the Soviet virologist Mikhail S. Balayan deliberately swallowed a pooled stool extract from affected patients, developed hepatitis, and then detected 27-to-30-nanometer virus particles in his own stool, demonstrating that a distinct, fecal-orally transmitted agent was responsible. (Such self-experimentation, while historically pivotal, would not meet today’s research-ethics standards.)
Together, hepatitis A through E illustrate a key and often-surprising fact: these viruses are not close relatives. They belong to entirely different virus families with different shapes, genomes, and routes of spread — A and E by contaminated food and water; B, C, and D mainly by blood and body fluids. They share only one thing, the organ they injure. “Hepatitis” names a target, the liver, far more than it names any single microbe — which is exactly why the disease’s causes had to be discovered one at a time.
Vaccines, Cures, and the Modern Legacy
The discoveries chronicled above were not ends in themselves; each became a tool that has saved enormous numbers of lives. Blumberg’s Australia antigen led first to blood-donor screening for hepatitis B and then to a hepatitis B vaccine — an early version derived from the surface antigen in human plasma, licensed in the United States in the early 1980s, followed by the modern, yeast-made recombinant vaccine that is now given routinely to infants worldwide. Because chronic hepatitis B is a leading cause of liver cancer, this was, in effect, one of the first vaccines able to prevent a human cancer. The identification of hepatitis A similarly yielded an effective inactivated HAV vaccine in the 1990s.
Hepatitis C followed the opposite but equally triumphant path. There is still no hepatitis C vaccine, but knowing the virus’s genome down to the molecule — the very work Houghton, Rice, and their colleagues began in 1989 — eventually enabled the design of direct-acting antiviral drugs. Beginning around 2014, these oral medicines turned a once-incurable, slowly fatal infection into one that can be cured in the great majority of patients with a few weeks of well-tolerated pills. Few transformations in the history of medicine have been so complete: a disease that long hid as an anonymous “non-A, non-B” menace in the blood supply is now both screenable and curable.
Looking back across the whole arc — from Hippocrates’ epidemic jaundice, through the false comfort of “catarrhal jaundice,” to Blumberg’s antigen and the cloning of hepatitis C — the history of hepatitis is a model of how medicine actually progresses. An ancient, visible symptom was slowly resolved into a set of distinct, invisible causes; wrong hypotheses were proposed and discarded; serendipity and dogged persistence each played their part; and the resulting knowledge was turned into tests, vaccines, and cures. The deeper detail on each virus — transmission, symptoms, diagnosis, and treatment — is taken up in the companion Hepatitis, Hepatitis B, and Hepatitis C articles.
Research Papers and References
The list below combines key peer-reviewed and historical sources on the discovery of the hepatitis viruses with curated PubMed topic-search links into the broader literature. Classical descriptions of epidemic jaundice (Hippocrates) and the nineteenth-century “catarrhal jaundice” idea (Virchow, 1865) are named in the article as historical sources rather than as modern citations. Each external link opens in a new tab.
- Blumberg BS, Alter HJ, Visnich S. A “new” antigen in leukemia sera. JAMA. 1965;191:541–546. (First report of the Australia antigen.) — PubMed: PMID 14239025
- Blumberg BS. Australia antigen and the biology of hepatitis B. (Nobel Lecture, 1976.) Science. 1977;197(4298):17–25. — doi:10.1126/science.325649
- Dane DS, Cameron CH, Briggs M. Virus-like particles in serum of patients with Australia-antigen-associated hepatitis. Lancet. 1970;1(7649):695–698. (The “Dane particle.”) — PubMed: PMID 4190997
- Feinstone SM, Kapikian AZ, Purcell RH. Hepatitis A: detection by immune electron microscopy of a viruslike antigen associated with acute illness. Science. 1973;182(4116):1026–1028. — doi:10.1126/science.182.4116.1026
- Choo QL, Kuo G, Weiner AJ, Overby LR, Bradley DW, Houghton M. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science. 1989;244(4902):359–362. (Cloning of hepatitis C.) — doi:10.1126/science.2523562
- Kuo G, Choo QL, Alter HJ, et al. An assay for circulating antibodies to a major etiologic virus of human non-A, non-B hepatitis. Science. 1989;244(4902):362–364. (First HCV antibody test.) — doi:10.1126/science.2496467
- Rizzetto M, Canese MG, Aricò S, et al. Immunofluorescence detection of a new antigen-antibody system (delta/anti-delta) associated to the hepatitis B virus in the liver and serum of HBsAg carriers. Gut. 1977;18(12):997–1003. (Discovery of hepatitis D.) — doi:10.1136/gut.18.12.997
- Balayan MS, Andjaparidze AG, Savinskaya SS, et al. Evidence for a virus in non-A, non-B hepatitis transmitted via the fecal-oral route. Intervirology. 1983;20(1):23–31. (Self-experiment confirming hepatitis E.) — doi:10.1159/000149370
- The Nobel Prize in Physiology or Medicine 2020 (Alter, Houghton, Rice) — for the discovery of the Hepatitis C virus — NobelPrize.org: 2020 Prize summary
- History and discovery of viral hepatitis — PubMed: history and discovery of viral hepatitis
- Blumberg, the Australia antigen, and hepatitis B — PubMed: Blumberg Australia antigen hepatitis B history
- Epidemic and “campaign” jaundice in history — PubMed: epidemic jaundice history
- MacCallum and the hepatitis A / hepatitis B nomenclature — PubMed: MacCallum hepatitis nomenclature
- Discovery of the hepatitis C virus (non-A, non-B hepatitis) — PubMed: hepatitis C virus discovery
External Authoritative Resources
- Nobel Prize — 2020 Physiology or Medicine (Hepatitis C virus)
- CDC — Viral Hepatitis
- World Health Organization — Hepatitis B fact sheet
Connections
- Infectious Disease
- Hepatitis (Overview)
- Hepatitis B
- Hepatitis C
- Cirrhosis
- Liver Disease
- All Conditions
- Jaundice — the visible sign that named this disease for centuries.